by Jim Steele
Is bleaching the legacy of a marvelous adaptation mechanism or a prelude to extirpation?
A Warm Evolutionary Legacy
Despite increasing confirmation of the Adaptive Bleaching Hypothesis and its ability to explain coral resilience, most people are unaware of its debate within the scientific community. The ability to rapidly adjust to changing environments by modifying their symbiotic partnerships has been the key to their success for millions of years. As one expert wrote, the “flexibility in coral–algal symbiosis is likely to be a principal factor underlying the evolutionary success of these organisms”.
Our modern day reef-building corals first evolved in exceedingly warm and stable climates when deep ocean temperatures were 10°C higher than today and palm trees dotted the Antarctic coast. As ice caps began to form in Antarctica ~35 million years ago sea levels fell and warm epi‑continental seas dried. After ocean depths had cooled for another 30 million years, Arctic ice caps began to form and the earth entered an age with multiple episodes of glacier advances and retreats causing sea levels to rise and fall. Just eighteen thousand years ago during the last glacial maximum, all our shallow reefs did not exist, as sea levels were 400 feet lower than today.
The 35 million year cooling trend increasingly restricted reef-building corals to more tropical latitudes where winter water temperatures remain above 16 to 18 °C. As their evolutionary history would predict, today’s greatest concentrations and greatest diversity of corals are found in the earth’s persistently warmer waters, like the Indo-Pacific Warm Pool. Likewise species inhabiting our warmest waters have undergone the fewest episodes of severe coral bleaching. Given their evolutionary history, coral’s greatest achievement has been enduring bouts of sustained climate cooling and rapid temperature swings. Even during warm interglacials coral battled cold temperatures dips. Studies of 7000-year-old fossil coral reefs in the South China Sea revealed high coral mortality every 50 years due to winter cooling events. Indeed most researchers believe past coral extinctions were most commonly due to cold events. Accordingly research has estimated that during the cold nadir of each ice age, coral reef extent was reduced by 80% and carbonate production was reduced by 73% relative to today.
As the last ice age ended, coral expanded their range with warming temperatures. At the peak of the Holocene Optimum 10,000 years BP (Before Present), coral adapted to tropical ocean temperatures in the heart of the Coral Triangle were 2.1 °C warmer than today. As illustrated above, temperatures cooled since then but frequently spiked or plummeted by 2 to 3 degrees over the course of a few centuries. One thousand years ago during the Medieval Warm Period, coral thrived in Pacific water masses that were ~0.65° warmer than in recent decades, then cooled ~0.9°C by the 1700s. Given coral’s evolutionary history, it is unlikely coral were better adapted to 1800s Little Ice Age temperatures versus Medieval Warm Period or 20th century temperatures. Emerging research now suggests coral bleaching has been an integral part of corals’ adjustment mechanisms to an ever-changing environment.
Coral Mortality and Resilience
There are 4 widespread misconceptions about bleaching propagated by tabloid media hyping climate doom and researchers like Hoegh-Guldberg. To clarify:
{kind=link}
- Bleaching is not always driven by warming temperatures
- Bleaching is not responsible for most coral mortality.
- Coral can rapidly respond to disturbances and replace lost cover within a decade or less.
- Bleaching, whether or not it results in coral mortality, is part of a natural selection process from which better-adapted populations emerge.
Multiple causes of bleaching
In contrast to researchers like Hoegh-Guldberg who emphasizes coral bleaching as a deadly product of global warming, bleaching is a visible stage in a complex set of acclimation mechanisms during which coral expel, shift and shuffle their symbionts, seeking the most beneficial partnership possible. Bleaching can be induced by stressful interactions between temperatures, disease, heavy rains, high irradiance from clear skies and competition with seaweeds. Indeed abrupt warm water events like El Nino have induced widespread bleaching and high mortality. But or of colder waters have also induced bleaching.In contrast to researchers like Hoegh-Guldberg who emphasizes coral bleaching as a deadly product of global warming, bleaching is a visible stage in a complex set of acclimation mechanisms during which coral expel, shift and shuffle their symbionts, seeking the most beneficial partnership possible. Bleaching can be induced by stressful interactions between temperatures, disease, heavy rains, high irradiance from clear skies and competition with seaweeds. Indeed abrupt warm water events like El Nino have induced widespread bleaching and high mortality. But cold winters or La Nina induced upwelling of colder waters have also induced bleaching.
NOAA has also contributed to these misconceptions by overemphasizing just warm-event bleaching. On NOAA‘s web page “What is Coral Bleaching”, NOAA reported, “the U.S. lost half of its coral reefs in the Caribbean” in one year due to warmer waters. But the Caribbean’s main cause of lost reefs was due to an outbreak of the White Band disease in 1981-82. White band specifically targets members of the genus Acropora, like the Staghorn and Elkhorn coral, reducing by 80% of their cover that once dominated the Caribbean reefs. However since the mid 80s experts reported coral cover has changed relatively little.
NOAA also downplayed cold temperature bleaching stating the 2010 cold event just “resulted in some coral death.” However NOAA’s statement stands in stark contrast to coral experts who reported the January 2010 cold snap was the worst coral bleaching and mortality event on record for Florida’s Reef Tract. They reported, “the mean percent coral mortality recorded for all species and subregions was 11.5% in the 2010 winter, compared to 0.5% recorded in the previous five summers, including years like 2005 where warm-water bleaching was prevalent.” Globally there has been an increase in observed cold bleaching events and 2010 was Florida’s first cold bleaching since the 1970s. Globally there have been several more reports of cold induced bleaching and then recovery as the waters warmed.
There is a perception that bleaching suddenly became more common only since the 1980s, leading some to speculate bleaching is due to rising CO2 and global warming. However, whether warming since the Little Ice Age is natural or anthropogenic, warming does not explain the increased observations of cold bleaching. More frequent observations of bleaching events may be partially due to the advent of remote sensing satellites that have allowed greater global coverage only since the 1980s. Furthermore determination of bleaching severity and mortality requires teams of divers to ground truth satellite data and fine-tune percentages of affected reefs. But SCUBA diving only became possible in the decades after Jacques Cousteau invented the Aqualung in the 1940s. Although natural rates of warming during the 30s and 40s were similar to today, coral reef studies were also hampered by the unsafe battleground between Japan and the Allies. War-time efforts such as the Battle of the Coral Sea, and fights to control the islands of Peleliu, Midway, Iwo Jima, the Philippines, or subsequent nuclear testing on the Bikini Atoll. The resulting reef devastation likely obscured any natural bleaching events.
We now know bleaching regularly happens due to seasonal fluctuations between high solar irradiance and warm temperatures of summer versus lower irradiance and cooler temperatures in winter. High irradiance can damage the corals’ symbiotic algae when photosynthesis runs too rapidly, while low irradiance detrimentally reduces photosynthetic output. Thus coral undergo natural adjustments to seasonal changes by expelling a portion of their symbiotic algae in summer. This leads to temporary or partial bleaching. Low light and colder temperatures slow photosynthesis, so coral increase their symbiont density in winter.
Similarly in response to changes in sunlight, the same species will alter their symbiotic partnerships as irradiance declines at increasing depths or when and where water turbidity alters irradiance. Bleaching is often temporary and mild as coral shuffle and switch their symbiotic algae in order to adapt, but sustained extremes, warm or cold, can prolong bleaching and starve the coral. Whether coral die or not depends on how quickly new symbionts are acquired relative to how much energy the coral has stored, or coral’s ability to feed on plankton as an alternative energy source.
All recent global bleaching events have been driven by El Nino events. The 1998 El Nino caused widespread mortality, an estimated 16% globally. Observed bleaching in response to warm tropical waters invading cooler regions aroused fears that climate change had contributed to this “unprecedented” event. However researchers have noted the relationship between warmer ocean temperatures and “bleaching has been equivocal and sometimes negative when the coolest regions were not in the analyses.” In other words coral living in the warmest waters were well acclimated to the warmest waters redistributed by an El Nino. Furthermore mortality did not always occur during periods with the warmest temperatures, but during the winter or ensuing cold La Nina conditions. Such observations suggest the rapid swings between anomalously warm El Nino and anomalously cold La Nina conditions are the most stressful.
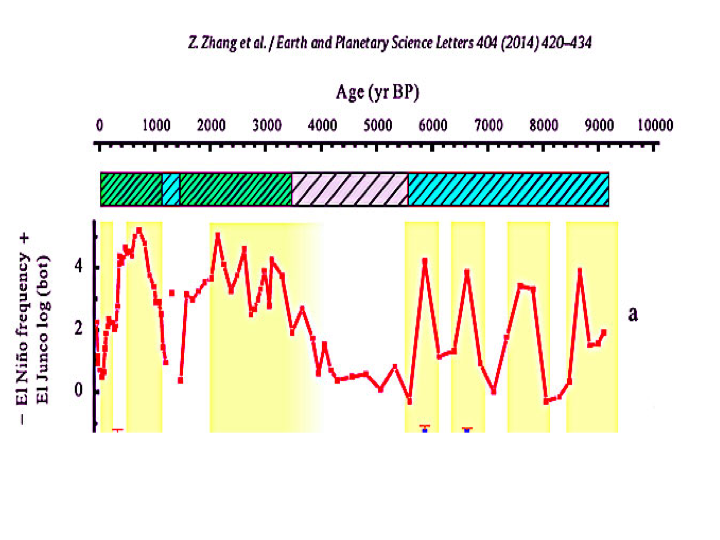
Stressful rapid temperature variations due to El Nino events have occurred throughout the past 10,000 years. As illustrated below from Zhang 2014, the frequency of El Ninos during the past century has been neither extremely high, nor extremely low. Most living coral species have survived over a million years of climate change and have endured the extreme El Nino frequencies of the past 3000 years including the Little Ice Age. El Nino events are a function of natural ocean variability and there is no consensus regards any effect from rising CO2 on El Nino frequency or intensity. To survive extremes from past natural variability, coral species had to be extremely resilient in ways that are just now being understood.Bleaching Causes the Least Mortality
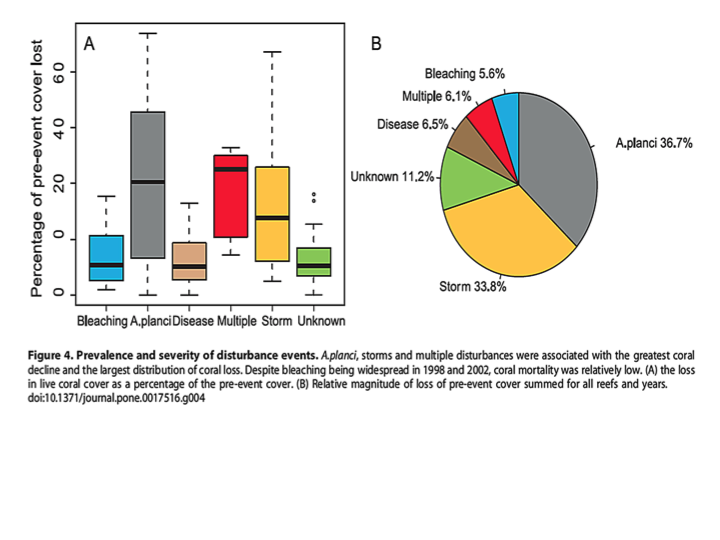
Most extreme bleaching events are associated with El Ninos, but the high mortality rates are not just a function of higher temperatures. Due to associated flooding and high rainfall, the resulting change in salinity disrupts coral osmosis, which can result in coral death. Furthermore tropical storms and heavy wave action are a major cause of lost coral reefs, but storms also bring heavy rains that also induce bleaching. Although some try to link storm-related mortality to climate change, there is no evidence of an increasing trend in tropical storms. As illustrated by the pie graph from Osborne 2011, in the Great Barrier Reef the explosion of the coral-eating Crown of Thorns starfish (A. planci) and tropical storms contributed to the greatest loss of coral colonies, 70.5%. Bleaching is a very minor contributor to coral mortality, just 5.6%, and that bleaching can be induced by warm or cold temperatures, heavy rains and floods or high irradiance from anomalously clear skies.Due to coral’s symbiotic efficiency and recycling of nutrients, corals dominate in nutrient-limited tropical waters. Normally those low nutrient conditions also prevent predators like the Crown of Thorns starfish (COTS) from rapidly reproducing because their plankton-feeding larvae typically starve. But increased inflow of nutrients due to landscape changes, agriculture run-off and sewage, has increased plankton blooms and thus the survivorship of COTS’ larvae. The ensuing population explosions of coral eating adults have decimated many reefs. COTS does not exist in the Caribbean. Instead coral there are battling bacterial diseases like white-band that can be spread by coral-eating snails. Humans have indeed tipped the balance in favor of COTS and in addition to destructive over fishing with dynamite and cyanide, those causes of coral death are the only factors we can remedy.
To understand coral resilience in the face of the variety of onslaughts, coral reefs must be seen as dynamic systems that oscillate over decadal periods, as well as centuries and millennia. Snapshots focused only on a few years when coral reefs decline misrepresents coral resilience and promotes false gloom and doom, as well as useless management plans. A long-term study of coral ecosystems of an island in French Polynesia demonstrates corals’ dynamics response to 32-years of storms, Crown of Thorns starfish and bleaching. Coral mortality is often measured as a function of the change in “coral cover”, and 45 to 50% of the healthy reef system around the island of Tiahura was covered with coral.
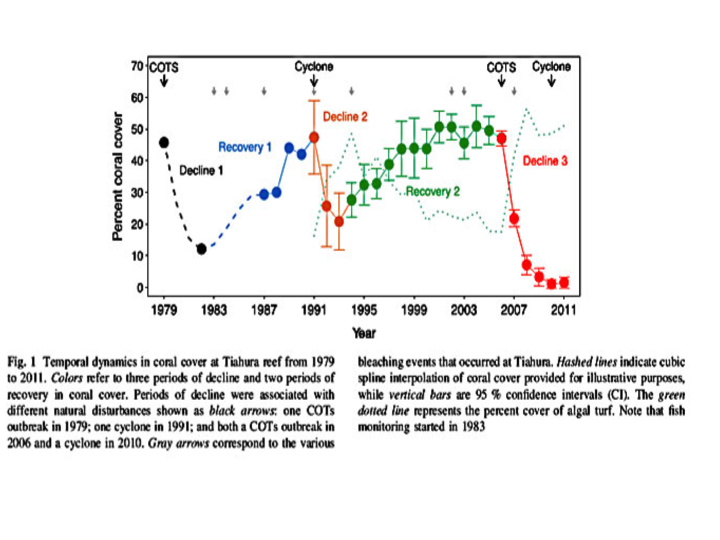
As illustrated below in Figure 1 from Lamy 2016, an outbreak of COTS removed 80% of the live coral cover between 1979 and 1982, reducing total coral cover to 10% of the reef. However by 1991 the coral had fully recovered. As designated by the small gray arrows at the top, three bleaching events occurred during that recovery period. Later destruction from a 1991 cyclone again reduced coral cover but again coral recovered reaching its greatest coverage of 50% by the year 2000. And again during that recovery there were 3 more bleaching events. Since 2006 the coral suffered their greatest loss due to another outbreak of COTS, quickly followed by another cyclone. High mortality promoted high seaweed cover (dotted green line) that has inhibited coral recovery. Over that time, coral bleaching was associated with periods of recovery, suggesting little if any detrimental effects. As will become clear shortly, one also could reasonably argue those bleaching events were beneficial..
Rapid Coral Recovery:
Tiahura’s coral recovery periods typically required 7 to ten years, and appeared to be unaffected by the 1998 El Nino. Several other studies have reported similar recovery periods, but some locations required 10 to 20 years to fully recover. In Australia’s Great Barrier Reef (GBF), the 1998 El Nino induced above average sea surface temperatures and salinity changes for 2 months triggering massive coral losses in the reef’s upper 20 meters. At the GBF’s Scott Reef, the upper 3 meters lost 80 to 90% of its living coral and the disappearance of half of the coral genera. Yet researchers observed, “within 12 years coral cover, recruitment, generic diversity, and community structure were again similar to the pre-bleaching years.” A similar long-term study in the Maldives observed a dramatic loss of coral during the 1998 El Nino but by 2013 the reefs also had returned to “pre-bleaching values”. Although a reef’s recovery sometime requires re-colonization by larvae from other reefs, a process known as re-sheeting or Phoenix effect can facilitate a reef’s speedy recovery. Often a small percentage of living “cryptic” polyps with a more resilient symbiotic partnership were embedded within a “dead” colony and survive extreme bleaching. They then multiply and rapidly “re-sheet” the colony’s skeletal remains.
In addition to rapid recovery of coral cover, researchers are finding bleached reefs have been increasingly less susceptible to subsequent bleaching. For example studies in Indonesian waters determined that two coral species, highly susceptible to bleaching, had experienced 94% and 87% colony deaths during the 1998 El Nino. Yet those same species were among the least susceptible to bleaching in the 2010 El Nino, with only 5% and 12% colony deaths despite a similar increase in water temperatures. Similarly, changes in resilience were observed in response to cold water bleaching in the Gulf of California. Increased resilience in response to a variety of bleaching events prompted the Adaptive Bleaching Hypothesis first proposed in 1993. The hypothesis suggests that although bleaching events are a response to stress, it creates the potential for coral to acquire totally new and different symbionts that are better suited to those stressful conditions. Contrary to Hoegh-Guldberg’s claim that coral reef systems will “experience near annual bleaching events that exceed the extent of the 1998 bleaching event by the year 2040”, scientists are increasingly observing the exact opposite. After reefs recover from severe bleaching, colonies have evolved enhanced resilience to future bleaching.
Coral Symbiosis, Symbiont Shuffling and Rapid Adaptation
A single coral colony is comprised of 100s to millions of individual “polyps” (seen above). Each polyp can be visualized as an upside down jellyfish (coral’s close cousins) with their backs cemented to a surface and tentacles extended outward to capture passing food particles, live prey, or new symbionts. However because coral live in nutrient depleted environments, in addition to filter feeding, polyps harbor single-celled photosynthesizing symbionts inside their cells. Those symbionts (aka zooxanthellae) typically provide ~90% of the coral’s energy needs. Just 40 years ago it was believed all corals were host to just one photosynthesizing symbiont, a single species from the dinoflagellate genus Symbiodinium. But thanks to technological advances in genetic sequencing, we now know a coral species can harbor several potential species or types of Symbiodinium algae, each capable of responding optimally to a different set of environmental conditions and coral physiology. As predicted by the adaptive bleaching hypothesis, improved genetic techniques have revealed a wondrously diverse community of symbionts that coral can choose from. Coral can no longer be viewed as organisms that only adapt slowly over evolutionary millennia via genetic mutation and natural selection. Coral must be seen as an “eco-species” (aka holobiont) that emerges from the synergy of the coral and its varied symbionts. And we now know those emergent eco-species can rapidly evolve with changing climates by shuffling and shifting those symbionts.
A single colony’s polyps are typically all clones resulting from asexual reproduction and on their own offer the colony scant genetic versatility. However within a colony, a wide variety of symbionts can be harbored within a small percentage of polyps, although one symbiont type typically dominates. That small percentage of “cryptic” polyps often survive severe bleaching episodes and then multiply rapidly over the skeletal remains in a process known as the Phoenix effect. Just one square centimeter of coral tissue typically harbors a million individual symbionts and on average those symbionts can double every 7 days. Thus after severe colony bleaching, a more resilient colony can arise in just a few years with better-adapted symbionts now dominating. Likewise symbiont variability within a reef results in some colonies bleaching while adjacent colonies of the same species do not. And similarly a varied symbiont and coral community allows neighboring reefs to adapt to their unique regional climates.
Figure 6 Colony on the left remains unbleached
Variations in coral reproduction can conserve an “ecospecies” or rapidly promote greater ecospecies diversity. Twenty-five percent of the coral species produce larvae inoculated directly from their parent’s symbionts. However 75% of the species produce larvae that initially lack a symbiont. Only after coral larvae settle on a surface, do those larvae engulf one or more different types of free-living Symbiodinium, drawing them inside their cells. As the larvae develop into mature polyps, coral typically keep the symbiont types best suited to the local microclimate and expel the others. In this manner completely new eco-species emerge.
Furthermore as conditions change, all species can shuffle their symbionts as polyps will expel their current residents and acquire a different type that had been harbored by a neighboring polyp. A colony can also shift its symbiont population by acquiring new types not yet hosted by the colony but are present in the reef. Due to improving genetic techniques, previously undetected types of symbionts with greater thermal tolerance are now being detected after bleaching events. Thus a combination of symbiont shuffling and shifting is the key to corals’ rapid adaptation. Although bleaching can result in coral death due to starvation when new symbionts are not acquired quickly enough, surviving polyps with their altered symbiont community have the potential to re-direct the reef on a trajectory that is better suited to the new environment. Or if conditions return to those prior to an extreme event, coral can re-acquire their old symbiont types.
Scientists have found that coral colonies nearer the surface often harbor a different type of symbiont than colonies living just a few meters deeper. The symbionts residing closer to the surface may be better adapted to high irradiance by making proteins that protect against too much ultra violet light or by modifying their photosystem. Conversely symbionts living at greater depths may photosynthesize more efficiently under low light conditions but are more susceptible to UV damage. Transplant experiments revealed that when coral colonies growing at greater depths were relocated closer to the surface, the polyps expelled their symbionts resulting in temporary bleaching. Bleaching allowed polyps to acquire new symbionts better adapted to higher irradiance. However colonies adapted to high-light surface conditions, photosynthesized much more slowly when transplanted to lower depths. Bleaching never happened and the coral died. Although experiments can force bleaching by raising temperatures, other controlled laboratory experiments found that in the absence of stress from high solar irradiance, anomalous temperatures 4 degrees above average still did not induce bleaching.
According to the adaptive bleaching hypothesis we can infer that bleaching events are not simply the result of recent global warming. Bleaching should have been ongoing for millions of years, as background temperatures have risen and fell. Thus we would expect that as the Little Ice Age ended and naturally temperatures rose, there should be observations of bleaching in the early 1900s. And indeed there are albeit limited. For example bleaching was reported in Florida on hot days in the early 1900s. But more telling, enough warm weather bleaching had been observed in the early 20th century that the Great Barrier Reef expedition of 1928-29 focused on warm weather coral bleaching when oceans were cooler than today and long before any possible CO2 warming effect.
Coral Response to Climate Change
Since his first Greenpeace-funded 1999 study, Hoegh-Guldberg has promoted catastrophic climate change as the biggest threat to coral reefs. His papers are frequently cited as evidence of climate related coral demise by some researchers and hyped by media outlets that boost readership by promoting climate catastrophes. The bases for his claims relied on 3 simplistic assumptions that a) bleaching is evidence that coral have reached their limit of maximum thermal tolerance, b) bleaching will increase due to global warming, and c) coral cannot adapt quickly enough to temperatures projected by climate models.
In 1999 Hoegh-Guldberg argued “thermal tolerances of reef-building corals will be exceeded within the next few decades” and coral reefs “could be eliminated from most areas by 2100” due to climate change. In his 2014 paper he continued to dismiss the emerging science supporting the adaptive bleaching hypothesis, belittling it as a “persistent mirage”. His catastrophic claims also intensified, suggesting “as much as 95% [of the world’s coral] may be in danger of being lost by mid-century.” To support his extirpation claim he cited two of his own previously published papers. Hoegh-Guldberg’s history of exaggeration and circular reasoning has led other coral experts to accuse him of “popularizing worst case scenarios”, while others have accused him of persistently misunderstanding and misrepresenting the adaptive bleaching hypothesis. Furthermore other researchers have pointed out the pitfalls and weaknesses in framing threats to coral based on a simplistic temperature threshold. They argue, “A view of coral reef ecosystems that emphasizes regional and historical variability and acclimation/adaptation to various environments is likely to be more accurate than one that sees them as characterized by stable and benign temperature regimes close to their upper thresholds.”
Improved genetic sequencing is increasingly providing evidence that in response to warm water bleaching events coral begin acquiring new heat resistant symbionts. The results below from Boulotte 2016 show that over the course of 2 years, colonies radically altered their symbionts. The pie charts represent the changing percentage of dominant symbiont types due to shuffling in a single reef species. The bar graphs list just the rarer symbionts and stars identify types not previously detected suggesting an ongoing shift. Symbionts “types” are characterized first by their genetic lineages known as clades. When the adaptive bleaching hypothesis was first proposed, only 4 clades were known. Now at least nine have been identified. The most heat resistant symbionts belong to clade D, but other heat resistant types have evolved within other clades. Many earlier acclimation studies simply identified a symbiont’s clade. But we now know each clade can harbor hundreds of types (potential species) and improved detection of those species is uncovering more shifting. The most heat resistant species identified to date belonged to clade C. As seen here, different types/species are identified as D_I:6 or D1.12. As illustrated below after 2 bleaching episodes, a new symbiont species from clade C began to dominate and previously undetected clade D symbionts began to appear more frequently in just 2 years.
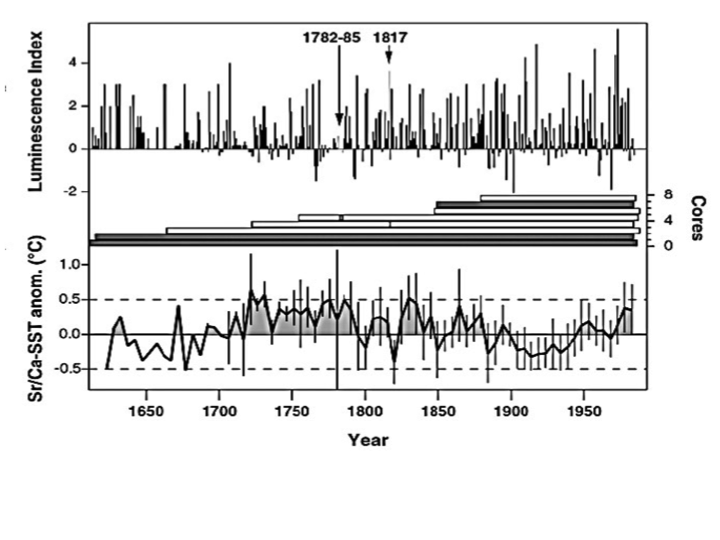
Nevertheless Hoegh-Guldberg 2014 continues to dismiss coral’s ability to rapidly adapt arguing, “current rates of change are unprecedented in the past 65 Ma [million years] if not 300 Ma.” But such exaggeration is pure nonsense. Ocean temperatures were warmer just 1000 years ago, and paleo-studies of temperatures in the Great Barrier Reef suggest local reef temperatures were higher between 1720 and 1820 as illustrated below from Hendy 2003. (Their luminescence index measures changes in salinity associated with monsoons). Perhaps CO2 concentrations are higher now than over the last 300 Ma. But given the extreme warmth just 65 million years ago, that is evidence that our climate is not very sensitive to CO2 concentrations, as realized by more researchers. In contrast to IPCC models that predict more warming that Hoegh-Guldberg ties to coral demise, climate experts note the Holocene temperature conundrum. While CO2 driven models simulate 6000 years of warming due to rising CO2, all the proxies indicate a cooling trend interrupted only by warming spikes.
Although coral genomes may evolve slowly, their symbionts have extremely fast generation times, averaging every 7 days. Furthermore the symbiont community consists of hundreds of symbionts that have already adapted to a wide variety of temperature, irradiance and salinity variables within different microclimates over the past million years. Symbiont shuffling and shifting is an evolutionary masterpiece that circumvents plodding evolutionary mechanisms of most organisms with long generation times and enables immediate adaptation.
A good summary statement is provided by Baker et al. “flexibility in coral–algal symbiosis is likely to be a principal factor underlying the evolutionary success of these organisms”.
Biosketch: Jim Steele is Director emeritus Sierra Nevada Field Campus, San Francisco State University and author of Landscapes & Cycles: An Environmentalist’s Journey to Climate Skepticism
JC note: This post was submitted via email, I did some minor editing on it. As with all guest posts, please keep your comments civil and relevant.Filed under: Adaptation, Oceans
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}