File attachments:
AttachmentSize
TPGVirusLikeNoOther.pdf2.84 MB
Language
English
0
No votes yet
Posted online July 12th 2017 LINK
Citation for this manuscript: Papadopulos-Eleopulos, E et al. HIV – A virus like no other. Posted at
the Perth Group website July 12th 2017. www.theperthgroup.com/HIV/TPGVirusLikeNoOther.pdf
No human experiment ought to continue if its scientific justification has been undermined.
Introduction
Richard Horton, Editor, The Lancet
What is now known as the Acquired Immune Deficiency Syndrome (AIDS) was first reported in 1981 as two diseases – a chronic pneumonia (PCP) caused by the fungal organism Pneumocystis carinii and Kaposi’s sarcoma (KS), a malignancy of uncertain histogenesis that principally involves the skin but may also occur in the gastrointestinal and respiratory tracts.1, 2 Neither PCP nor KS were new diseases. What was new was the exponentially escalating incidence of what were two formerly rare diseases and their proclivity for a minor subset of young, sexually promiscuous, drug using homosexual men.3 Subsequently more diseases were added under the umbrella term “AIDS indicator diseases” which currently number 29, including PCP, KS, tuberculosis,4 candida (yeast) infections, lymphoma and cervical cancer.
Among the first to put forward a theory to account for the high frequency of KS and PCP in homosexual men were researchers belonging to what Luc Montagnier, from the Pasteur Institute in Paris, calls the “Retrovirology Club”.5 During the 1970s the Retrovirology Club tried to prove, albeit unsuccessfully, that cancer is caused by viruses.6 Since KS is a malignancy, retrovirologists, in particular Robert Gallo from the US National Institutes of Health, proposed a viral theory of AIDS. The observations the viral theory was intended to explain were threefold: the high frequency of KS; a few opportunistic infections, principally PCP; and a decrease in a cell type, the T4 (CD4) lymphocyte, in the peripheral blood of the homosexual patients. Subsequently the theory was also claimed to explain opportunistic infections and T4 cell decrease in intravenous drug users and haemophiliacs.
It was accepted that no single infectious agent could directly cause the heterogeneous group of AIDS “indicator” diseases. Hence, it was proposed that viral-induced destruction of T4 cells (acquired immune deficiency), the "hallmark" of HIV infection, inevitably led to the appearance of KS and the opportunistic infections.7 In other words, viral infectionT4 cell destructionthe clinical syndrome (AIDS). The virus, now known as the human immunodeficiency virus, was said to be transmitted principally via sexual intercourse, blood and blood products. Further information on immune deficiency HERE
Origins
The first report of an “AIDS virus” was in a May 20th 1983 paper published in Science by scientists at the Pasteur Institute led by Luc Montagnier. They claimed to have isolated a retrovirus, lymphadenopathy-associated virus (LAV), from a homosexual patient code name BRU who was at risk of AIDS and had a pre-AIDS prodrome.8 In May 1984 scientists at the National Institutes of Health in the USA led by Robert Gallo published four papers also in Science in which they claimed to have isolated a retrovirus, human T-cell lymphotropic virus-III (HTLV-III), from 26/72 patients
with AIDS and concluded their data “suggest that HTLV-III [HIV] may be the primary cause of 1
AIDS”.9-12 In 1986 Gallo recounted his 1984 data as “The results presented in our four papers provided clearcut evidence that the aetiology of AIDS and ARC [AIDS-related complex, another prodrome] was the new lymphotropic retrovirus, HTLV-III”.13 The latest (2015) 19th edition of Harrison’s Principles of Internal Medicine asserts, “In 1983, human immunodeficiency virus (HIV) was isolated from a patient with lymphadenopathy, and by 1984 it was demonstrated clearly to be the causative agent of AIDS”. In three papers published in 1984,14-16 Gallo and his associates were the first to claim characterisation of the HIV genome and as a result were the first to introduce its use into clinical practice. By 1986 LAV and HTLV-III were accepted to be the same virus and Montagnier’s and Gallo’s viruses were renamed human immunodeficiency virus (HIV).17 As Joseph Sonnabend, an infectious disease specialist physician practising in New York City at the beginning of the AIDS era, summed up the zeitgeist, “Very early on in the epidemic, before HIV was discovered, there were two theories [for AIDS], one was that there was a new agent out there, and the other was the multifactorial [lifestyle] theory...so there is a competition between two theories and different interests latch on to different theories for different reasons...The conservative family values lobby liked the single virus because it says if you have sex outside marriage you could die. If you're a gay man – die. The gay leadership liked it too so they were joining hands with their enemies in a sense, both favouring the single virus theory. It takes the view away from lifestyle and puts it on a single virus”.18
The scientific and medical communities readily opted for the retroviral theory and rapidly adopted the belief that the apparent spread of “HIV” represented a global health emergency, with “real, and potentially significant, risks to national, regional, and global security from the pandemic”;19 and for over three decades have resisted every alternative view. In 2008 Montagnier and Barré-Sinoussi were awarded the Nobel Prize in Physiology or Medicine “for their discovery of human immunodeficiency virus".20 On 20th May 2016 the Pasteur Institute tweeted “33 years ago today Françoise Barré-Sinoussi and Luc Montagnier published in the journal Science the discovery of the retrovirus that causes AIDS”.21 However, according to Anders Vahlne, Professor in Clinical Virology, Karolinska Institute, Stockholm, “In reality, in my view there is no evidence whatsoever in this [Montagnier’s 1983] paper that a new human retrovirus has been isolated!”22, 23 The question arises, does the evidence presented in Montagnier’s 1983 paper prove the existence of HIV? If not, did Montagnier prove its existence in a subsequent paper? Or have other scientists published such evidence?
Viruses and proof for their existence
A virus is a microscopic, infectious particle. Infectious refers to the cycles of transmission and replication whose steps include release of viral particles from infected cells, their entry into uninfected cells (transmission), intracellular synthesis of particle proteins and nucleic acid, terminating in assembly and release of new viral particles. HIV is said to belong to the Family Retroviridae which have RNA genomes and according to the theory of retroviruses, an additional step in their replication cycle is the reverse transcription of their genome. In other words, the synthesis of a DNA copy of their RNA genome using an enzyme called reverse transcriptase. After copying, viral DNA is integrated into the host cell DNA as the “provirus”. Virologists refer to the subsequent synthesis of new viral RNA and proteins, the assembly and release of particles, as “expression” of the proviral genome.
All virologists including retrovirologists and in particular those who gave the world the human immunodeficiency virus – Luc Montagnier, Francoise Barré-Sinoussi, Jean Claude Chermann and Robert Gallo – acknowledge that to prove the existence of a virus one must purify the virus particles.24, 25 Purification is required for several reasons, including the following:
1. Viruses replicate only in living cells. Since cells and viruses are composed of the same biochemical constituents, separation of particles from cellular material is essential for defining which nucleic acid and proteins belong to the virus particles.
2
2. To prove the particles are infectious. In other words, it is particles, not other factors, that are responsible for the production of new particles. This requires purification of both sets of particles.
3. To demonstrate their biological and pathological effects. 4. To obtain antigens (proteins) and nucleic acids for use in antibody and genomic tests
(including “viral load”26) respectively.
The method used to purify retroviral particles is based on the opportune fact that such particles have a buoyant density of 1.16 g/ml in a sucrose solution. This property enables their separation from cellular material using a procedure known as density gradient ultracentrifugation. If a cell culture is producing a retrovirus the viral particles are released from infected cells into the culture fluids. The purification procedure begins by placing an aliquot of culture supernatant on top of a sucrose solution prepared such that its density gradually increases from the top to the bottom of a test-tube (see diagram below). The tube is spun at high speeds generating an enormous force that propels the supernatant constituents down through the density gradient towards the bottom of the tube. Over several hours each constituent reaches a place in the gradient where its density is equal to that of the surrounding solution, whereupon it stops sedimenting. In this manner the constituents become trapped (concentrated) in several density regions (“bands”) according to their differing buoyancies. At the completion of the procedure the centrifuge is stopped, the tube removed, its base punctured and aliquots of fluid, effectively the individual density bands, are sequentially removed. The 1.16 g/ml band is collected for electron microscopic [EM] and biochemical analyses.
Density gradient centrifugation
In the five 1983/84 Science papers, Montagnier and Gallo and their colleagues claimed to have purified HIV using density gradient centrifugation, “characterised” (identified) the HIV particle proteins, showed that the particles are infectious and proved (Gallo) HIV the cause of AIDS. The 1.16 g/ml density band material was declared to be “purified”, “pure”, retrovirus particles, despite the fact neither group published electron microscopic images to substantiate this claim. Neither have they nor any other scientist since published such images.
Nevertheless, HIV protagonists accept that Montagnier and his colleagues were the first to prove the existence of HIV, as documented in their 1983 Science paper “Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS)”; although their
3
paper concluded “The role of this virus in the etiology of AIDS remains to be determined”. Indeed, this is the research which led to Montagnier’s and Barré-Sinoussi’s Nobel prize.
One year later, using similar methods, Gallo and his colleagues repeated Montagnier’s experiments and claimed to have proven HIV is the cause of AIDS. By December 1984, they also claimed to have characterised the HIV genome. Hence, if there is an AIDS-causing retrovirus, the evidence in these 1983/84 publications should unambiguously confirm its existence. This being the case, in order to avoid any charge of misinterpretation, we will describe each of Montagnier’s experiments, with his interpretation followed by our commentary. The latter will include a general discussion of Montagnier’s and Gallo’s virus experiments, and a description of the series of scientific errors which we claim led to the construction of the “HIV” hypothesis. Following this, Gallo and his colleagues’ genomic data will be examined in detail.
Before proceeding with the description and analysis of these experiments it is necessary to clarify the terms “virus isolation” and “virus purification”. Montagnier, Gallo and many other scientists frequently invoke these terms in support of their claim to have proven the existence and characterisation of HIV. To the non-scientist and layman, “isolation” and “purification” are the same: isolating, that is, separating an object from all other different objects, defines the process of purification. However, in virology these terms are not synonymous.27-31 When the Canadian documentarian Brent Leung asked UK retrovirologist Robin Weiss for an explanation he replied, “Isolation and purification are jargon words in virology...they mean different things to different people...they’re not very precise”.32 Yet isolation and purification are the basis of the peer- reviewed, "clear-cut, exhaustive and unambiguous" evidence said to prove the existence of HIV and its causative role in AIDS.33-35 The fact is that in virology, while purification retains its everyday meaning, “isolation” is an expediential term virologists assign to data they claim are proof a particular virus exists.36
4
Montagnier’s isolation experiments
In his 2012 documentary House of Numbers? Brent Leung interviewed Montagnier and Barré- Sinoussi:
Leung: What is the purpose of the purification? 32 Montagnier: To make sure you have a real virus.
Leung: Going back to 1983, when trying to prove the existence of a new virus, why was purification important? Barré-Sinoussi: It was important to prepare kits for antibody detection. OK? Because we wanted these diagnosis kits to be as specific as possible. If you use a preparation of virus which is not purified of course you will detect antibody to everything, not only against the virus but also against all the proteins that are produced in the supernatant...Now when this virus [HIV] is in this [cell culture] supernatant it’s not purified. OK? Because the cells are releasing plenty of things, not only the virus...cellular proteins...so on, OK?...so that means in the supernatant you have a mixture of everything, including the virus. Then you have to purify it...OK...this is the second step...then you try to purify the virus from all this mess.
The first two experiments, involving the enzyme reverse transcriptase, incorporate an error that was to become ubiquitous in “HIV” research.
First Montagnier experiment
Method: Montagnier cultured T-lymphocytes obtained from a lymph node excised from the patient BRU. BRU “was a 33-year-old homosexual male who sought medical consultation in December 1982 for cervical lymphadenopathy and asthenia...Examination showed axillary and inguinal lymphadenopathies. Neither fever nor recent loss of weight were noted. The patient had a history of several episodes of gonorrhea and had been treated for syphilis in September 1982. During interviews he indicated that he had had more than 50 sexual partners per year and had traveled to many countries”.
5
To the culture of BRU’s lymphocytes a number of chemicals were added including the mitogen phytohaemagglutinin (PHA). “Samples were regularly taken for assay of reverse transcriptase and for examination in the electron microscope”. Result: Detection of reverse transcriptase (RT) activity in the culture. No EM images published. Montagnier’s interpretation: “Virus production”. BRU infected with a retrovirus.
Comments
In 1971 Nobel laureate Howard Temin, the discoverer of reverse transcriptase, reported the isolation of a reverse transcriptase from uninfected rat cells and concluded that reverse transcriptase activity does not “necessarily represent oncogenic [retro] viruses”.37 In 1976 none other than Gallo showed that reverse transcription occurs in normal, non-virus infected, PHA stimulated cells.38 Mitogenic stimulation (with PHA or other mitogens) is obligatory in HIV “isolation” experiments: the phenomena claimed to represent HIV isolation do not appear without mitogenic stimulation. Several common microbes, including bacteria39 and hepatitis B virus (a common infection in AIDS patients, including their T4 cells40), reverse transcribe. The current list of over one hundred, retro-transcribing (reverse transcribing) viruses is HERE
In House of Numbers, Nobel laureate David Baltimore told Leung, “reverse transcription is very widespread”.41 In Australia in 2001 the non-specificity of RT was even publicised in a popular share trading magazine article evaluating the investment potential of biotechnology companies.42 Yet the HIV/AIDS scientific literature is replete with claims of detection, transmission, isolation and even quantification of HIV based on nothing more than the detection of reverse transcriptase activity.43, 44 As late as 1997 Jaap Goudsmit, one of the best known HIV experts, asserted, “The BRU lymph node was first cultured in early January 1983 and, on January 15, it shed an enzyme absolutely unique to the lentivirus [retrovirus] group”45 (emphasis added). Despite all the evidence to the contrary (much of it their own from the 1970s) leading HIV experts still claim reverse transcriptase is retroviral specific.
Indeed, when interviewed by the French investigative journalist/documentarian Djamel Tahi, Montagnier's co-worker Jean-Claude Chermann said: “The second point is related to the detection of RT activity which is a retrovirus specific enzyme” (D. Tahi, personal communication). As recently as 2006 the HIV expert David Ho told a PBS interviewer, “Reverse transcriptase is an enzyme of retroviruses...one way to look for retroviruses is simply to measure reverse transcription capability, and in fact that is how Barré-Sinoussi and her colleagues discovered HIV...They showed this reverse transcriptase activity as being transmissible in the tissue culture”.46
For Montagnier detection of reverse transcriptase activity ≡ HIV infection of BRU. The same conclusion is asserted by all other HIV researchers including Gallo and his colleagues in 1984 performing similar experiments on their patients. However, the interpretation RT activity ≡ retrovirus is contradicted by the scientific evidence. When interviewed in July 1997 at the Pasteur Institute by Tahi, Montagnier correctly referred to “RT activity, which is the enzyme characteristic of retroviruses”.47 He did not say specific “of retroviruses”.
All Montagnier’s experiments were performed using either invalid controls or no controls. A control is an essential part of a scientifically valid experiment designed to show that the factor being tested is actually responsible for the effect observed. In the control experiment all factors, apart from the one under test, are exactly the same as in the test experiments, and all the same measurements are carried out. The use of controls is elementary and in the case of retroviruses, crucial. In Montagnier’s experiments controls are mandated to ensure phenomena interpreted as “retroviral”, such as reverse transcriptase activity, are not the result of unforeseen, confounding non-retroviral factors or the expression of endogenous retroviruses48 which are present “in all of us”.49, 50 (“Endogenous retrovirus” signifies the presence of sequences resembling retroviral genomes in human DNA which are not expressed as infectious particles (hence the term is a misnomer). 8% of the human genome is said to consist of such sequences.49, 50. “The production of endogenous
6
viruses in cell cultures can start spontaneously or can be induced by chemical agents or radiation”51).
It is never possible to specify every confounding factor in an experiment but at least the design of controls must account for every factor that is known. These include the in vivo physiological state of patients from whom the putatively infected cells and sera are obtained; and the in vitro conditions under which the cells are cultured, manipulated and maintained. There are many scientific publications documenting that non-retrovirus-infected cells, cultured under the same conditions as “infected cells”, produce one or more of the same phenomena said to prove retroviral “isolation” from “infected” cells. In 1976 the retrovirologist George Todaro asserted that failure to produce retroviral-like particles in cell cultures “may reflect the limitations of in vitro cocultivation techniques",52 that is, limited by the conditions prevailing in a particular cell culture at the time that experiment is performed. Before the AIDS era Gallo, Weiss and other scientists published papers showing that “The expression of endogenous retroviruses can affect the results of seemingly unrelated experiments”.38, 53
Controls used in HIV research must be cells and sera obtained from patients who are as similar as possible to AIDS patients but who do not have AIDS or belong to an AIDS risk group. The similarity must encompass the clinical, haematological, biochemical, serological (hypergammaglobulinaemia) and metabolic (cellular oxidation) findings that are well documented in AIDS patients. The control experiments should be run in parallel with the test experiments, with both test and controls treated in exactly the same manner. To minimise bias the experimenter must be blinded as to which subjects are the test group and which are the controls. No such data were reported by Montagnier in any of his experiments. Failure to use valid controls, or even any controls, pervades HIV research. Montagnier’s first “control” consisted of a lymphocyte culture from a healthy individual in which RT activity was not detected. This “control” was invalid, because regardless of exposure to a putative virus, such cells would not be in a condition comparable to those from a patient with AIDS.
Montagnier did not have proof the RT activity was due to a retroviral enzyme. Furthermore, he had no proof the enzyme was reverse transcribing a retroviral RNA or even a cellular RNA. He detected RT activity by introducing an artificial RNA into the culture, an RNA to which was attached a short segment of artificial DNA. This RNA-DNA is known as a template-primer and consists of a 100-200 sequence of the same ribose nucleotide sequence (the template), primed at one end with a single DNA nucleotide sequence. The artificial template-primer used by Montagnier, ubiquitous in HIV research, is An.dT12-15, [aka ((rA)n.(dT)12-15) and An.dT15]. This template-primer is transcribed not only by reverse transcriptase but by all cellular DNA polymerases. Montagnier knew that in 1980 there was proof that “Among a number of template primers, (rA)n.(dT)12-18 has been most frequently employed since RT shows high activity with this template●primer. However, the cellular DNA polymerases (pol β and pol γ) also effectively utilize the same template●primer”.54 In fact, in 1975 one of these polymerases, DNA polymerase γ, was defined as the cellular enzyme that “copies An.dT15 with high efficiency but does not copy DNA well”.55 The latter is confirmed in a review of DNA polymerase γ published by Laurie Kaguni from the Department of Biochemistry and Molecular Biology, Michigan State University, in 2004.56 A more detailed account of reverse transcription and reverse transcriptases is HERE
7
Second Montagnier experiment
Method: BRU’s T-cells were co-cultured with healthy blood donor T-cells. Result: Detection of RT activity. No EM data published. Montagnier’s interpretation: “Propagation” (transmission) and “isolation” of a retrovirus.
Comment
Proof of transmission requires the introduction of purified, retrovirus-like particles into an uninfected cell culture followed by the appearance of particles morphologically and biochemically identical to those added and with negative results in controls. Despite reporting “Samples [of the culture supernatant] were regularly taken for...examination in the electron microscope”, Montagnier did not publish evidence for the existence of virus-like particles in his co-culture. Montagnier had no control. The second control should have consisted of healthy donor lymphocytes co-cultured with T-lymphocytes from a sick individual as defined above. Even if RT activity were specific to retroviruses, the detection of RT activity cannot be considered proof of transmission. The RT activity could have been confined to the BRU cells, as it was in the first experiment. Proof of transmission requires proof the RT activity was produced by the healthy blood donor T-cells. In this experiment there was no such evidence. Again, even if the enzyme were retroviral specific, reverse transcriptase activity is not isolation of a retrovirus. Detection of cardiac or hepatic enzyme activity in the blood of a patient does not mean the laboratory has isolated the patient’s heart or liver.
8
Third Montagnier experiment
This illustrates the second major error in the construction of the “AIDS virus”, this time involving purported viral particles. Following the failure to report particles in the culture from the first two experiments, supernatant obtained from the second experiment was incubated with lymphocytes obtained from umbilical cord blood of two placentas.
Method: Supernatant from the BRU + healthy blood donor T-cell co-culture was added to umbilical cord T-cell cultures. Result: One electron micrograph of the culture showing retrovirus-like particles. Montagnier’s interpretation: BRU infected with “a typical type-C” retrovirus.
Comment
Montagnier’s single electron micrograph from the umbilical cord lymphocyte culture is the only electron microscopic evidence that BRU’s cells were infected with a retrovirus57 but this claim is surrounded by many uncertainties.
In 2010 Barré-Sinoussi described the events leading up to the electron micrograph:32, 41
...and then [after finding RT activity in the cell culture] ...we immediately call our guy who was responsible for electron microscopy and said please, could you look under the microscope, whether you can see virus particle, and if it resemble to a retrovirus...and after, after, quite, it was very difficult because it was only few cells infected, so it was a very difficult task, for him, to find the cells that was just producing these particles but, finally he found it, and he found one lymphocyte, with a budding particle, typical of retrovirus, and, very close from this cell, one complete mature particle that resembled to a retrovirus. (In 2005 Montagnier’s electron microscopist and co-author Charles Dauguet told Djamel Tahi he was asked to examine the cell culture only after spending 15 days in an unsuccessful search for particles in density gradient “purified virus” material58).
Montagnier had no control. The control for experiment three, the third control, should have been the addition of supernatant from the second control experiment (as defined above) to the umbilical cord lymphocyte culture. The culture as well as density gradient purified supernatant from test and
9
controls should have been submitted for electron microscopy with Dauguet expending the same time and effort examining both sets of samples. That there were no controls was confirmed by Dauguet when he was asked if he examined controls: “No...I do not think so. The samples on which I worked were from infected cultures”.58
By definition retroviruses are “enveloped viruses with a diameter of 100-120 nm budding at cellular membranes. Cell released virions [cell free particles] contain condensed inner bodies (cores) and are studded with projections (spikes, knobs)”.59 According to their mode of assembly and fine structure they are divided into sub-families and genera. To date, neither Montagnier nor Gallo has published an electron micrograph of particles claimed to be “HIV” showing all the morphological characteristics of retroviral particles.
Montagnier et al EM of “virus-producing cord lymphocytes”
In 1984 Montagnier’s particles were reported as “typical type-C” retroviral particles, a genus belonging to the Oncovirinae Subfamily of Retroviridae.60 A year later Gallo also reported his HIV as type-C particles. Then also in 1984, HIV experts (including Montagnier) reported HIV a member of another Oncovirinae genus, that is, type-D retroviral particles.61, 62 In 2003, using atomic force electron microscopy (with resolution in fractions of a nanometre), Yuri Kuznetsov and his colleagues showed that HIV particles “are virtually indistinguishable from virions [virus particles] of MuLV” (murine leukaemia virus).63 MuLV is the prototype type-C retrovirus particle.64 Then in 1986, when the names LAV and HTLV-III were dropped, Montagnier’s and Gallo’s type-C particles were renamed HIV and classified a lentivirus, a genus belonging to another Subfamily of Retroviridae, that is, Lentivirinae.17 (The particles that Montagnier displayed as HIV in his Nobel lecture65 in December 2008 defy classification66, 67 HERE).
The retroviral taxonomy in use prior to and during the 1980s is described in Perspectives in Medical Virology Volume 3 dated 1987, “Criteria for classification of retroviruses are predominantly morphological features as seen in ultrathin sections: site of core assembly (preformed in the cytoplasm or formed during the budding process at the plasma membrane); shape and size of surface protrusions (spike- or knob-like); presence or absence of electron-lucent space between envelope and core in immature particles, and shape and position of cores in mature particles”.60 Cognisant of these facts it is highly unlikely electron microscopists would have mistakenly classified the same virus a member of two Subfamilies and three genera of the Family
Retroviridae. Either there is no agreement as to which Subfamily or genus “HIV” belongs, or, since 10
the Montagnier and Gallo viruses are “typical type-C” particles and type-C viruses are not lentiviruses, the retrovirus now called HIV cannot be what Montagnier discovered and Gallo reported in 1983 and 1984 respectively.
Hans Gelderblom’s model of the “ideal” HIV particle.59, 68
HIV experts agree there are spikes/knobs on the surface of the viral particle whose presence is an absolute requirement for infectivity.69 The knobs are said to consist of two proteins, gp120 and gp41 (gp = glycoprotein). Hans Gelderblom from the Robert Koch Institute in Berlin is the leading expert on HIV electron microscopy. In 1987 he and his colleagues published a model of the “ideal” HIV particle, claiming that “On the ‘ideal’, intact HIV particle 72 knobs can be determined”. However, in their most detailed electron microscopic studies Gelderblom’s group reported that
11
“cell-released, ‘mature’ HIV particles represent by far the majority of virus structures...The loss of surface knobs apparently correlates morphologically with virus maturation. Immature and/or budding HIV particles are “spiked”, but they are rarely observed” and are seen only “on metabolically impaired cells”.59, 68, 70 Immature particles have knobs but by definition are not infectious. Since the knobs are crucial for infectivity but are lost during maturation, the mature particles cannot be infectious either.
In 2006 Ping Zhu and colleagues71 published a paper in Nature “Distribution and three-dimensional structure of AIDS virus envelope spikes”. Despite the title suggesting a discourse on HIV “envelope spikes”, the fact is they analysed and "generated a three-dimensional (3D) model of the SIV [simian immunodeficiency virus] Env [envelope] spike", not HIV. Furthermore, they claimed the HIV particles have “14 ± 7 Env spikes per particle (range 4 to 35) (see examples in fig. 2b-d)”. However, fig. 2b-d shows only “surface-rendered models” of HIV virions with “presumptive Env- spikes” (emphasis added). In the images in fig. 1b (HIV-1 below) which presumably are their best “Examples of putative Env spikes on selected virions”, it is difficult, if not impossible, to see any spikes on the HIV-1 particles (emphasis added). The HIV-1 image also contains structures resembling “putative Env spikes” in parts of the image where there are no particles. These are opinions shared by disinterested scientists competent in the field.72
The Zhu caption reads: “Figure 1 | Representative tomographic images of mutant SIV and wild- type HIV-1....Examples of putative Env spikes on selected virions are indicated by arrowheads...Scale bars, 100 nm” (top right in each image).
This is consistent with the researchers’ previous work where “Immunoelectron microscopic analysis using sera from HIV-1-infected patients showed little labeling [by antibodies that bind to the knob/spike proteins] of mature HIV-1 particles”73 and with Hans Gelderblom’s findings. The differences between Gelderblom and Zhu et al are: (i) Gelderblom claims the spikes are rapidly lost in the process of maturation while in Zhu and colleagues’ view the spikes are not lost but their number is determined by the low incorporation of the HIV surface proteins into the particles to begin with; (ii) in Gelderblom's view “it was possible that structures resembling knobs might be observed even when there was no gp120 [spikes] present, i.e. false positives”,74 while Zhu et al call them “putative Env spikes”. In the last sentence of his Nature commentary75 on the Zhu et al paper Dennis Burton made reference to the 2003 Journal of Virology paper by Kuznetsov et al63 remarking that “Atomic force microscopy studies have given a different view of the HIV envelope spike”. In their paper Kuznetsov et al explain that they chose atomic force microscopy for their analysis because “Cryo-electron microscopy does, however, still suffer from problems in
12
interpretation due to superposition of features”. Their data showed that “The clusters of gp120 do not form spikes on the surface of HIV as is commonly described in the literature" and they "found no evidence that the gp120 monomers form threefold symmetric trimers...We suggest that the spikes observed by negative-staining electron microscopy may be an artifact of the penetration of heavy metal stain between envelope proteins. Indeed, the term “spike” appears to have assumed a rather imprecise, possibly misleading definition, and might best be used with caution”. In other words, the “different view of the HIV envelope spike” Burton attributes to Kuznetsov is that there are no HIV envelope spikes.
Taxonomy, and the absence of spikes/knobs, are not the only “HIV particle” problems. Some of the others include:
1. Type-C retroviral-like particles are ubiquitous. They are present “in the majority, if not all, human placentas”76 as well as in tissues from fish, snakes, pheasant, quail, partridge, turkey, tree mice, agouti [rodents], tapeworms, insects and mammals.77 The electron micrograph of Montagnier’s “HIV” particles was that of a culture of lymphocytes obtained from umbilical cord blood of two human placentas.
- In Montagnier’s third experiment, cell-free supernatant from a BRU cell culture was added to cord blood lymphocytes. Hence, the particles must have arisen from these cells. However, cord blood lymphocytes express budding virus-like particles “independently of HIV infection”.78-80 Since Montagnier had no controls it is impossible to attribute the type-C particles observed in the third experiment to transmission of a retrovirus originating in BRU.
- Cells from AIDS patients are frequently co-cultured with immortal cell lines such as H9 which is claimed to facilitate what Gallo calls “continuous production” and “true isolation”81 of HIV. However, electron microscopy reveals a plethora of retroviral-like and non- retroviral-like particles in these cultures.59, 82, 83, 84 HIV experts have maintained a long silence on the origin, nature, role and relationship of these particles to AIDS.85
-
Particles identified as HIV are observed in the enlarged lymph nodes of AIDS patients. However, identical particles are also observed with the same frequency in enlarged lymph nodes of patients who do not have AIDS and are not at risk of AIDS. Such was reported in an extensive, detailed and blinded electron microscopic study reported by O'Hara and
13
colleagues from Harvard. "HIV particles" were found in 18/20 (90%) of patients with enlarged lymph nodes attributed to AIDS whereas identical particles were found in 13/15 (87%) of patients with enlarged lymph nodes not attributed to AIDS.86
O’Hara et al
5. According to the HIV experts David Ho87 and Xiping Wei,88 HIV positive individuals have massive HIV infection from inception in whom an “estimated average total HIV-1 production was 10.3 x 10(9) [109] virions per day”. Wei cites Michael Piatak that “Virtually all HIV-1- infected individuals, regardless of clinical stage, exhibit persistent plasma viraemia in the range of 102 to 107 virions per ml”.89 Gelderblom writes “preparations for electron microscopic diagnostic procedures require particle concentrations of 106 to 108/mL. Therefore, negative evidence is not an absolute diagnosis. A number of effective concentration or immunologic procedures exist that markedly increase sensitivity of electron microscopic diagnostics for samples with lower particle concentrations”. Such methods can increase sensitivity 5-1000 fold.90 In 2014 it was reported that recently infected patients may have viraemia as high as 108 particles per ml.91 This being the case it would undoubtedly be possible to confirm viraemia using electron microscopy. Yet, to date there is not one published electron micrograph proving the presence of retroviral particles in any patient with “HIV viraemia” including “in the range of 102 to 107 virions per ml”.26
Many images purporting to be “HIV” are artists’ or computer graphics, not original, untouched electron micrographs. For example, the front page of the International AIDS Society Newsletter, March 2007, is entitled “AIDS Denialists. This edition’s feature article examines the global impact of AIDS denialism”. About 75% of the page is occupied by a multi-coloured picture apparently meant to represent part of a cell with budding and cell free “HIV” particles. The picture is not an electron micrograph but a computer graphic with a caption “Image: HIV daughter cells bud off the surface of a T-cell”.92 However, viruses cannot be “daughter cells” because viruses are not cells.
Another example is a news item published in Nature on November 20th 2003 entitled “Medical journal [British Medical Journal] under attack as dissenters seize AIDS platform”.93 This one page article includes a scanning electron micrograph with a caption which reads “The BMJ’s website carries postings that deny that HIV, seen here in a white blood cell, causes AIDS”. The image,
14
resembling a spoonful of spaghetti, occupies about a quarter of the available space, presumably reflecting its importance. However, the source of the electron micrograph is not given, the “HIV” is unlabelled and has no size bar. The appearance of the cell in the micrograph is unlike any white blood cell that has ever traversed the vascular system. If the particles displayed are indeed a retrovirus they are obviously on the cell and not in the cell as the author claims. Moreover, these surface particles are cylindrical, not spherical, and are several microns in length. Such appearances and dimensions would be not only unique to “HIV” but to any other retrovirus seen using electron microscopy in any situation. Our group wrote to Nature questioning this uncharacteristic lack of scientific rigour and suggested that because of the importance of this matter Nature could either seek clarification from the HIV experts or preferably arrange a scientific debate between the two sides adjudicated by disinterested scientists. In this manner the matter could be resolved once and for all. Our letter was rejected but we were told that Nature “will probably publish a correction”. Although we are regular readers of Nature we have yet to see one.
Caption: “The BMJ’s website carries postings that deny that HIV, seen here in a white blood cell, causes AIDS”
The fact is that as with reverse transcriptase activity, retrovirus-like particles are non-specific. Retrovirus-like particles can be detected in individuals with non-AIDS-related illnesses and even no illness.69, 94, 95 Nonspecific findings are common in medicine. Fever, for example, is a feature of hundreds of diseases and not a diagnosis. Just as fever indicates that a disease is present but does not specify which disease, retroviral-like particles and reverse transcriptase activity are indicative of a possible disorder, but are not retroviral-specific.
15
Fourth Montagnier experiment
This experiment and its interpretation by Montagnier and his team provided the basis for the third major error at the foundations of “HIV/AIDS” science. It enabled both Montagnier and Gallo to give to the world “HIV” diagnostic kits which to this day have never been validated.
Method: Umbilical cord lymphocytes “infected” as per the third experiment were incubated for 20 hours with radio-labelled [35S] methionine. (Methionine is an amino acid incorporated into proteins produced in the culture. Its radioactivity enables their detection following exposure to a photographic plate). From a sample of the cell free supernatant “The virus was purified by banding on a sucrose [density] gradient”. In the “purified, [radioactively] labeled virus” (that is, the 1.16 g/ml band), RT activity was detected. Then, serum from (a) BRU; (b) a second patient; (c) two healthy individuals; and (d) antibodies directed against the p24 protein of HTLV-I were added to the “purified, labeled virus”.
Result: A reaction between antibodies present in the BRU serum and three proteins (p25, p45, p80) in the “purified, labeled virus” but no reactions with HTLV-I, the second patient and the healthy donor sera. Montagnier’s interpretation: BRU infected with a new retrovirus. The p25 protein is a constituent of the new virus. The p45 protein is not a retroviral protein because “The 45K protein may be due to contamination of the virus by cellular actin”8 (the molecular weight of actin is 41K). p80 was not further mentioned but in a later paper Montagnier claimed it, like the p45 protein, is cellular.96 The BRU virus is new because the 1.16 g/ml band material did not react with an antibody to the HTLV-I p24 protein. (Note: Montagnier’s p25 is now known as p24). LINK to the BRU protein data.
Comments
Montagnier and his colleagues did not publish evidence that their 1.16 g/ml band, the band which they claimed to be “purified, [radioactively] labeled virus”, contained particles with the morphological characteristics of a retrovirus, pure or impure, or indeed any particles of any kind. They only showed that in the band they could detect:
(a) RT activity;
16
(b) proteins which reacted with antibodies present in BRU’s serum.
However, since many proteins including reverse transcribing enzymes, either free or embodied in particles other than retroviruses (cellular debris, viruses) also band at 1.16 g/ml, and reverse transcription is not specific to retroviruses, finding reverse transcriptase activity in the 1.16 g/ml band is not proof for the existence of retrovirus particles, much less purified retrovirus particles.
Patients with AIDS and those at risk, as typified by BRU, have an abundance of antibodies, including auto-antibodies (antibodies directed against self-constituents). Montagnier himself showed that AIDS patients and those at risk have antibodies to the two ubiquitous self-proteins actin and myosin.97 The concentration of antibodies in HIV/AIDS patients is typically 70% higher than in normal individuals, including autoantibodies. In fact, individuals with AIDS, AIDS-related complex and those at risk have an ever increasing list of autoantibodies: circulating immune complexes, rheumatoid factor, anti-cardiolipin, anti-nuclear factor, anti-cellular, anti-platelet, anti-red cell, anti-actin, anti-DNA, anti-tubulin, anti-thyroglobulin, anti-albumin, anti-myosin, anti-trinitrophenyl, anti-thymosin, anti-interleukin and anti-lymphocyte antibodies.98-100
In the Tahi interview Montagnier accepted that AIDS patients have a plethora of antibodies “but antibodies are very specific. They know how to distinguish one molecule in one million”.101 Even if Montagnier were correct, at best all one can conclude from a reaction between an unknown protein X in the 1.16 g/ml band and an unknown antibody Y in BRU’s serum is that BRU has been exposed to X. But from this reaction it is impossible to determine the identity or origin of X or Y. Even if the origin of X or Y were known, neither can one determine the origin of the other. This is because an antibody does not react exclusively with the antigen that induced its appearance.102 Antibodies induced by and directed against a given protein may react with other proteins, sometimes many proteins. Immunologists define these reactions as “cross-reactions” or “cross- reactivities”. The prevalence of cross-reactions is increased in patients who have elevated levels of antibodies, such as HIV positive and AIDS patients. In other words, far from distinguishing “one molecule in one million”, antibodies are promiscuous, a behaviour that “shocked” the immunological community and led to their use of this descriptor.103 The scientific literature is replete with data showing antibodies are not “very specific”, are not “razor sharp”104 and cannot “distinguish one molecule in one million”.105, 106 This is why an antibody test to diagnose infection with a particular agent should not be introduced into clinical practice before that agent has been used as the gold standard comparator to prove the test is specific for that agent. In the case of the HIV antibody tests this most basic requirement has not been met.100, 107-119
Montagnier knew BRU had a surfeit of antibodies but not the origin of any. The data from his fourth experiment were that the 1.16 g/ml band contained three proteins which reacted with antibodies present in BRU. Even if one assumed that a known protein (p25/24) causes the appearance of only one antibody which reacts with it and no other substance, the best one could say from Montagnier’s evidence is that at some stage of his life BRU came into contact with that protein. But nothing can be said about the origin of p24 itself. Since BRU’s serum contained a multitude of antibodies and antibodies cross-react, any of his antibodies could have reacted with any of the proteins present in the 1.16 g/ml band (including p24), even if BRU had never come in contact with any of them. Yet, from such a reaction Montagnier claimed to have determined the origin of both the protein and the antibody – a scientific impossibility.
In this experiment more than in any other, controls are of fundamental importance because the results were interpreted as proving the existence of a new retrovirus, HIV, and its proteins. And the same “HIV” proteins were soon to be incorporated as antigens in antibody test kits for the widespread diagnosis of HIV infection.110 (The commonly used tests are methodologically different but detect the same antibodies – the enzyme immunoassay (EIA) (aka the Enzyme Linked Immunosorbent Assay, ELISA) and Western blot. In the EIA the patient’s serum is added to a mixture of “HIV” proteins. In the Western blot the same proteins are separated along the length of
17
a nitrocellulose strip so that individual antibody/protein reactions ("bands") can be seen and interpreted).
Again Montagnier had no control. In this fourth experiment Montagnier should have had two 1.16 g/ml bands. One obtained from the supernatant of the umbilical cord lymphocyte test culture in experiment three (the “purified, labeled virus” band) and one, the fourth control, from the supernatant of the third control experiment. Both bands should have been tested with the BRU serum and serum from AIDS patients and those at risk, as well as control sera obtained from sick individuals as defined above. Since AIDS patients and those at risk have hypergammaglobulinaemia120 and are oxidised, and since oxidation leads to increased levels of autoantibodies and their “unmasking” with their “growing list of specificities”,121-123 the control sera must have the same properties. To the “purified, labeled virus”, Montagnier added sera from (a) BRU; (b) a second patient; (c) two healthy individuals; and (d) a serum containing antibodies directed against the p24 protein of HTLV-I. The only reactions were between BRU serum and three proteins including p24.
Montagnier concluded that BRU’s antibodies “recognised”124 a p24 protein in the “purified, labeled virus” material and hence proved p24 is an “HIV” protein. However, if a p24 protein had also been “recognised” by control sera in the “purified, labeled virus”, or if p24 had been found where it should not have been, that is, in the control 1.16 g/ml band, then it would have been impossible for Montagnier to reach this conclusion. In fact, from the time antibody testing began there were studies (and many more since) which show that reactivity to the BRU p24 “HIV” protein is prevalent worldwide amongst individuals who do not have AIDS and are not at risk of AIDS, including healthy individuals. So much so, that by 1987 these data necessitated a redefinition of the criteria for interpreting the “confirmatory” HIV Western blot test.125 Initially, a positive Western blot was a reaction with p24 or p41 or both but subsequently reactivities to several other “HIV” proteins were added. It must be stressed that prior to 1987 reactivity solely to p24 was considered a positive antibody test and proof of infection regardless of reactivity with other “HIV” proteins.126
There are many examples that affirm reactivity to p24 by a wide range of sera. Perhaps because “HIV” was thought to originate in Africa, as soon as antibody tests became available Montagnier and Gallo were the first among many to conduct such tests in Africans.
- In a 1984 study from Kinshasa “Prevalence of antibodies to lymphadenopathy-associated retrovirus [HIV] in African patients with AIDS”, Montagnier and 19 colleagues reported reactivity to p24 in 6/26 (23%) of controls.127
-
In a study in the same year entitled “Evidence for exposure to HTLV-III in Uganda before 1973” Gallo tested stored blood collected between August 1972 and July 1973 from 75 healthy, six- year old Ugandan children. 50/75 (67%) were HIV-positive.128 Since the HIV theory of AIDS requires mother-to-child transmission as the cause of HIV antibodies in children this age, and since African AIDS is purportedly spread by heterosexual intercourse, Gallo expected seropositivity in children to be mirrored by their parents. Even today in Africa, barrier contraception is problematic, so by 1984, AIDS in Uganda should have been commonplace if the antibody tests are proof of HIV infection, and HIV causes AIDS. At that time 50 per cent of antibody-positive homosexual men in the West were developing AIDS within 10 years, and the figure was considered to be 1-2 years shorter in poorer countries. Before 1997, untreated AIDS was generally fatal within one to two years. By 1997, when antiretroviral therapies were introduced in the West, the Ugandan population should have been in obvious and serious decline.129 Yet, the first AIDS case in Uganda was not diagnosed until 1984,130 that is, at least 15 years after these healthy, six year old children acquired their “HIV” infection; while between 1980-88 the population growth rate in Uganda increased from 3-3.5% per annum and since then has averaged 3.4%,131 three times greater than the United States and the United Kingdom.
18
- “25% of a sample of hospital workers in Zaire were seropositive in 1984".132
- "15.5% of blood donors were found to be positive at Kigali in Rwanda in 1984".133
- "41 out of 410 (10%) of healthy medical personnel from Mulago Hospital Kampala were positive for HTLV-III/LAV. Five out of 30 (17%) of controls outside the hospital were positive. Four out of 10 (40%) of control patients deemed sexually immature were also found positive".134
- In 1989 Joan Genesca and her colleagues conducted Western Blot assays in 100 EIA negative samples from healthy blood donors; 20 were found to have HIV bands which did not fulfil the then (1989) criteria used by the blood banks for a positive WB. These were considered indeterminate WB, (WBi), with p24 being the predominant band (70% of cases). Among recipients of WBi blood, 36% were WBi 6 months after transfusion but so were 42% of individuals who received WB negative blood. Both donors and recipients of blood remained healthy. They concluded that WBi patterns "are exceedingly common in randomly selected donors and recipients and such patterns do not correlate with the presence of HIV-1 or the transmission of HIV-1".135 Genesca also noted that “48-64% of donors repeatedly reactive for anti-HIV-1 by EIA have WBi patterns. The frequency of such patterns in low-risk populations is so high as to suggest that, as with EIA, most such reactions represent false-positive results”.
- In 1988 Annamari Ranki and her colleagues reported antibodies that reacted with HIV “core proteins ” (p24 and p55) in non-HIV-infected homosexual men, as well as individuals with cutaneous T-cell lymphoma or prodrome, dermatological diseases and multiple sclerosis.136
- In 1992 Felix Agbalika and his colleagues reported that “Abbott HIV-1-EIA kits detected non- neutralizable antigens in early post-transplantation sera from 12 kidney, five bone-marrow and two heart recipients. Using in-house immunoblots [Western blots] prepared from positive non- neutralizing antigen sera, a 25-30 kD protein was detected and shown to be the cause of the false HIV antigen cross-reactivity”.137
- In 1994 Oscar Kashala and Myron Essex reported antibody test data on leprosy patients and their contacts (“family members and other persons living within 1.6 km of the leprosarium who interacted daily with the patients”). Leprosy is caused by Mycobacterium leprae, a bacterium that “shares several antigenic determinants with other mycobacterial species, including M. tu- berculosis”. They reported reactivity with p24 in 24/39 (61.5%) contacts.138
- Lundberg and his colleagues from the US Consortium for Retrovirus Serology Standardization reported that 127/1306 (10%) of individuals at "low risk" for AIDS including "specimens from blood donor centers" had a positive HIV Western blot by the "most stringent" of the US criteria, that is, the presence of antibodies to p24, p32 and gp41 or gp120/160.139
- In 1992, Jorg Shüpbach, the principal author of the third and co-author of the fourth of the 1984 papers published by Gallo's group on HIV isolation, reported that the whole blood cultures of 49/60 (82%) of "presumably uninfected but serologically indeterminate individuals and 5/5 sero- negative blood donors were found positive for p24".140
- According to the AID vaccine Clinical Trials Group "The presence of p24 band was common among low-risk, uninfected volunteers and complicated the interpretation of the Western blot test results".141
- The "HIV proteins (p17, p24)" appear in the blood of patients (previously negative for all HIV markers) following "transfusions of HIV-negative blood and UV-irradiation of the autoblood".142
-
According to Fauci “By definition, Western blot patterns of reactivity that do not fall into the positive or negative categories are considered "indeterminate””. Furthermore, “There are two possible explanations for an indeterminate Western blot result. The most likely explanation in a low-risk individual is that the patient being tested has antibodies that cross-react with one of the proteins of HIV. The most common patterns of cross-reactivity are antibodies that react with p24 and/or p55”.143
19
- Sera from dogs “recognise” the “HIV” p24 (and other “HIV” proteins). In 1991 Strandstrom and colleagues reported that 72/144 (50%) canine blood samples "obtained from the Veterinary Medical Teaching Hospital, University of California, Davis" tested in commercial Western blot assays, "reacted with one or more HIV recombinant proteins [gp120–21.5%, gp41–23%, p31– 22%, p24–43%]".144
- In 1990 Michael St. Louis and his colleagues anonymously tested 89,547 blood specimens from 26 US hospital patients. This study not only excluded patients in the known AIDS risk groups but also patients with almost a hundred other diagnoses including "gunshot and knife wounds” all of which pose meagre if any risk of HIV/AIDS.145 They reported that 0.7% - 21.7% of men and 0-7.8% of women aged 25-44 years were EIA and HIV Western blot positive,146 that is, they had antibodies that reacted with many of the “HIV” proteins including p24.
- If as Montagnier states, “antibodies are very specific. They know how to distinguish one molecule in one million. There is a very great affinity...With monoclonal antibodies you fish out really ONE protein”47 (emphasis in original) or, as Gustav Nossal asserts, monoclonal antibodies are “razor sharp”,147 then a monoclonal antibody directed against the “HIV” p24 protein should react with material from tissues where a retrovirus HIV is present and never with anything else. However, in 1997 Achim Kramer published a paper “Molecular basis for the binding promiscuity of an anti-p24 (HIV-1) monoclonal antibody”148 with evidence that a monoclonal antibody directed against the “HIV” p24 protein “recognises” proteins found in bacteria, yeasts, amoebae, rabbits, monkeys and non-HIV-infected humans. The fungi include Candida albicans, the agent that causes one of the common AIDS indicator diseases.
Montagnier’s evidence in regard to the discovery of HIV has been disputed by at least three scientists, Anders Vahlne,22 Professor in Clinical Virology, Karolinska Institute, Stockholm, Sweden; the well-known HIV expert Jaap Goudsmit, principal researcher of the Amsterdam Cohort Studies on HIV infection and AIDS among homosexual men and HIV drug users45; and Udaykumar Ranga from the Jawaharlal Nehru University, New Delhi.149
Vahlne wrote, “Regarding whom should get the credit for the discovery of HIV, this review should enable the reader to come to his or her own conclusion. Mine, however, is different from that of those of my fellow faculty members that presently make up the Nobel Committee for the Nobel Prize in Physiology or Medicine...In reality, in my view there is no evidence whatsoever in this [Montagnier’s 1983] paper that a new human retrovirus has been isolated!”22, 23 Vahlne went on to argue that "With the data presented, the virus they isolated could well have been HTLV-I or in particular HTLV-II...However, the proof that a new human retrovirus (HIV-1) was the cause of AIDS was first established in four publications by Gallo's group in the May 4th issue of Science in 1984”. Vahlne’s interpretation is problematic for many other reasons. Discussion HERE
In his paper “The Saga of the HIV Controversy Nobel Prize in Physiology or Medicine – 2008” Ranga argued “‘The general understanding’ today is that while Montagnier was the first to isolate the AIDS virus, it was Gallo who ‘accomplished’ to establish the cause and effect relationship between the virus and AIDS. This statement and many others that reinforce this notion are far from the truth. The single publication of Montagnier in 1983 and all the publications of Gallo in 1984 collectively did NOT establish that AIDS was caused by their viruses” 149 (emphasis in original).
Let us give the last word to Montagnier and his electron microscopist colleague and co-author Charles Dauguet. Fourteen years after he claimed to have proven the existence of a new and unique retrovirus HIV by purifying the viral particles Tahi asked Montagnier:
Tahi: Why do the EM photographs published by you, come from the culture and not from the purification? [the 1.16 g/ml density band] Montagnier: We saw some particles [in the 1.16 g/ml band] but they did not have the morphology typical of retroviruses. They were very different.
20
Tahi: Why no purification? Montagnier: I repeat we did not purify.
Tahi (to Dauguet): How long have you searched in purified gradients before finding the first images of the virus? Dauguet: I first worked on gradients of purified virus for 15 days. Tahi: Have you found viral particles?
Dauguet: We have never seen virus particles in the purified virus. What we have seen all the time was cellular debris, no virus particles (D. Tahi, personal communication).
Tahi (to Montagnier): Gallo did it [purified]? Montagnier: Gallo? I don’t know if he really purified. I don’t believe so. I believe he launched very quickly into the molecular part, that’s to say cloning.
Tahi: Today, are the problems about mass production of the virus, purification, EM pictures at 1.16, resolved? Montagnier: Yes, of course. Tahi: Do EM pictures of HIV from the purification exist?
Montagnier: Yes. of course. Tahi: Have they been published? Montagnier: I couldn't tell you...we have some somewhere...but it is not of interest, not of any interest.47, 150
In other words, Montagnier and his colleagues had a purified 1.16 g/ml band, but what they purified was cellular debris, not retrovirus particles. Nonetheless, they called the 1.16 g/ml band “purified, labeled virus”.
Since both the Pasteur Institute electron microscopist and Montagnier admit that the 1.16 g/ml band contained cellular debris and no retrovirus-like particles, it follows that all proteins in the band, including that responsible for the reverse transcriptase activity as well as p24, must be cellular proteins. This means there are no retroviral proteins and thus there can be no retrovirus
21
RNA and no retrovirus HIV. Yet, for 34 years Montagnier’s p24 has been considered the most specific HIV protein and its detection considered not only proof for infection but also used to quantify HIV and prove its “isolation” in cultures by the use of an antibody reaction. The latter was the proof of “Reduction of maternal-infant transmission of human immunodeficiency virus type 1 with zidovudine treatment”, reported in the Pediatric AIDS Clinical Trials Group 076 study.151
HIV ISOLATION POST-MONTAGNIER
Robert Gallo and his associates were the first to assert that Montagnier’s “Isolation of a T- lymphotropic retrovirus” from BRU was not “true isolation”. They subsequently claimed that it was their four, May 1984 Science papers that contain the proof of “true isolation”. However, there are very few differences between the Montagnier and Gallo isolation experiments.
Montagnier had only the one patient, BRU, and used an umbilical cord lymphocyte culture in attempts to prove the existence of viral proteins and thus the existence of LAV (HIV). Gallo had 72 patients and instead of using umbilical cord T-cells as medium for growing the purported virus, he used the H9 clone of a “T-cell line...termed HT...derived from an adult with lymphoid leukemia”.81, 152 The H9 cell line was cultured with tissue originating from patients with AIDS or those at risk of AIDS. The supernatant of the H9 cell culture was banded in a sucrose density gradient and the 1.16 g/ml band, that is, the material Gallo called “purified virus”, was incubated with sera obtained from many AIDS patients and those at risk.
Unlike Montagnier’s one patient, BRU, whose antibodies reacted with three proteins in the “purified, labeled virus”, Gallo had many patients whose sera contained antibodies which reacted with either a p41 or a p24 or both proteins present in his “purified virus”. Unlike Montagnier, Gallo claimed both proteins were viral but regarded p41 more specific than p24. Gallo like Montagnier found many proteins present in the cells from his cultures that also reacted with antibodies in his patients’ sera but unlike Montagnier, he claimed these proteins were either viral or virally induced. Montagnier’s proof for transmission and isolation was detection of RT activity in two consecutive cultures. Gallo’s defined the criteria for "Detection and isolation of HTLV-III [HIV] from patients with AIDS and pre-AIDS” as:
"Samples exhibiting more than one of the following were considered positive”: 1. “Repeated detection of a Mg2+ -dependent reverse transcriptase activity in supernatant
fluids”;
- “Virus observed by electron microscopy”;
-
“Intracellular expression of virus-related antigens detected with antibodies from seropositive
donors or with rabbit antiserum to HTLV-III”; -
“Transmission of particles, detected by RT assays or by electron microscopic observation,
to fresh human cord blood, bone marrow, or peripheral blood T lymphocytes".9
Detection is not isolation. And no matter what Gallo meant by isolation, his criteria cannot be used to prove isolation. RT activity, even if detected in a thousand serial cultures is not proof for isolation. Observing “virus” by EM does not prove the particles detected are a virus, much less “HIV”, or that the “virus” has been isolated. These criteria may be used for detection but if and only if there is proof each is 100% HIV specific.153 Such proof requires purification of the virus particles. Gallo did not publish such proof in 1984 nor has he since.
Even if “more than one” of Gallo’s criteria were proof of “Detection and isolation”, his results would still be highly problematic – a fact Gallo acknowledged. Gallo tested 72 AIDS patients but “Detection and isolation” was positive in only 26/72 (36%). This means that nearly two thirds (64%) of his AIDS patients were not infected with “HIV, the virus that causes AIDS”. Gallo “solved” this problem in a highly implausible manner, as he explained to Huw Christie, editor of Continuum Magazine, during an interview conducted in 1998 at the Geneva International AIDS Conference. Gallo told Christie, “Sometimes we had Western blot positive but we couldn’t isolate the virus. So we got worried and felt we were getting false positives sometimes so we added the Western blot.
22
That’s all I can tell you. It was an experimental tool when we added it, and for us it worked well ‘cos we could isolate the virus when we did it”.154, 155 It may have “worked” for Gallo but it is scientifically invalid. A Western blot is another technology for detecting an antigen/antibody reaction and, like his criterion (3), is not virus isolation. An antibody/antigen reaction can be used for detection if and only if the reaction has been proven specific, proof of which first requires purification of the virus.
In his book Science Fictions John Crewdson describes how Gallo claimed his “rabbit antiserum to HTLV-III” was “among his most important contributions. ‘It wasn’t until the rabbit antibody that we knew we had the cause of AIDS’”.156 To produce rabbit antibodies directed against HIV one must inject rabbits with purified HIV particles or proteins. The question is, how did Gallo possess an antiserum to prove “Detection and isolation” of a retrovirus before he had proof for the “Detection and isolation” of the same virus? If Gallo’s antiserum was obtained by injecting rabbits with the material in the 1.16 g/ml density band his rabbits would have produced antibodies that reacted with possibly everything in that material, no matter what its origin. But since Gallo like Montagnier did not publish even one electron micrograph157 to show the “purified virus” contained retrovirus particles, much less purified particles, it is likely that the “purified virus”, contained nothing but cellular debris. This means that Gallo’s “rabbit antiserum to HTLV-III” was nothing more than rabbit antibodies to an unknown number of cellular proteins.
Similarly, Gallo claimed he employed “antibodies from seropositive donors” to detect “Intracellular expression of virus-related antigens [proteins]”. However, in order to determine which donors are seropositive [have antibodies that react with the “intracellular antigens”], one must first obtain the “virus-related antigens”, which can only be obtained by purifying the virus.
Gallo judges his laboratory’s introduction of the H9 cell line a highly significant factor in his successful “true isolation” and characterisation of HIV. Reading the 1984 Science papers one gets the impression the HT cell line from which his colleague Mikulas Popovic developed the H9 clone originated in Gallo’s laboratory.152 However, the National Institutes of Health enquiry into Gallo’s laboratory practices158 established that HT is nothing but HUT-78, a cell line developed in another laboratory in the late 1970s by Adi Gazdar. Publishing in Nature in April 1983 Gallo reported that HUT-78 cells “contained HTLV[-I] proviral [retroviral] sequences”.159 In 1991 and 1993 Robert Dourmashkin and his colleagues reported that H9 cell cultures produce retrovirus-like particles when not infected with HIV.78, 79
23
Dourmashkin et al
Caption: “Electron micrographs of selected membrane regions of fixed and sectioned cells showing cell-associated VLP [virus-like particles] budding from uninfected H9 cells” (emphasis added).
Furthermore, although Gallo considered his immortal H9 cell line of utmost significance in the isolation of HIV, Montagnier’s view is that in immortal cell lines “It is a real soup” of retrovirus particles.47 Gallo, like Montagnier, did not use valid controls. Gallo’s “true isolation” was no more “true” than Montagnier’s.
In 2003 the Perth Group emailed Gallo asking if he was aware of the Tahi interview and Montagnier’s admission there were no electron micrographs of the BRU "purified virus". Did clinicians have cause for concern about the obvious implication of Montagnier’s answer? Had clinicians spent two decades diagnosing patients with a non-existent HIV? Gallo replied “Montagnier subsequently published pictures of purified HIV as, of course we did in our first papers. You have no need of worry. The evidence is obvious and overwhelming”. In fact there was not a single electron micrograph of purified “HIV” published by Gallo in 1984 or since, or by Montagnier.157
1997: THE “PURIFIED VIRUS” IS REVEALED
In March 1997 Pablo Gluschankof, the leader of a large European HIV research collaborative, published a paper in Virology which began by acknowledging that HIV "used for biochemical [RNA/DNA] and serological analyses [antibody and antigen tests] or as an immunogen [injecting the material claimed to be the virus or virus proteins into animals to produce antibodies] is frequently prepared by centrifugation through sucrose gradients", and that in none of the studies "has the purity of the virus preparation been verified".160, 161
This means that for 14 years after the existence of HIV was accepted as proven, the community of HIV experts was using proteins and RNA as reagents for HIV diagnosis (antigen, antibody and nucleic acid tests), monitoring and research without proof the proteins and RNA originated in a viral particle. The true nature of “purified virus” came to light only when Gluschankof and a second group from the US National Cancer Institute led by Julian Bess162 each published electron- microscopic images – two density gradient purified culture supernatants from “HIV-infected” cultures and one from non-infected cultures. Even a cursory inspection makes it plain that whatever material is portrayed in these images, it is not pure.
24
“Purified virus” – Gluschankof et al
The caption to the Gluschankof EMs reads: “Purified HIV-1 preparations are contaminated by cellular vesicles. Purified vesicles from infected H9 cells (a) and activated PBMC (b) ...or from noninfected H9 cells (c) Virions are indicated by arrows” (PBMC = peripheral blood mononuclear cells; underlining in text and boxes surrounding particles in (c) ours).
Comments
- The material in electron micrographs (a) and (b) originated from “infected” cultures. Being density gradient purified material it should consist of nothing but purified retrovirus particles yet all three micrographs are labelled “Purified [cellular] vesicles”.
- The average diameter of the five particles indicated by arrows as “Virions” is 140nM. This exceeds the taxonomically defined diameter of retroviral particles (100-120nM in the 1980s and later revised to 80-100nM163). Furthermore, the five “Virions” lack the cone-shaped cores and lateral bodies required by lentiviral morphology. None have the spikes/knobs which are deemed an absolute requirement for infectivity (see above).
-
In electron micrograph (c), the “Purified vesicles”, noninfected material, there are particles resembling the “HIV” “Virions” in (a) and (b). In the Bess electron micrographs (not shown) the objects designated HIV have an average diameter of 234 nM and none has a diameter less than 160 nM. On this basis they cannot be a retrovirus.164 When we emailed Bess he agreed the particles are this size but could not provide an explanation. He said he would consult with his electron microscopists and let us know the outcome. Unfortunately he did not.
25
- Naturally scientists are motivated to present their best results, and the best electron micrographs from both groups show that in reality the “purified virus” consists of a variegated collection of cellular microvesicles165, 166 and other debris of various shapes and sizes none of which is a particle bearing the complete set of morphological features required by lentiviruses. Not even the particle dimension designated by The International Committee on Taxonomy of Viruses is correct.
-
The Bess paper included a protein electrophoresis167 of “HIV-infected” and noninfected density gradient purified material. If the “HIV-infected” material contains a retrovirus HIV as well as cellular material (microvesicles) then, compared to the non-infected material, it must contain the extra 15 proteins said to constitute the HIV virions. However, the Bess data show there are no extra proteins. Apart from quantitative differences in three of the proteins which Bess labelled p6/7, p17 and p24 in the “HIV-infected” material (B&C), the protein profiles of B&C and the uninfected preparations (A) are identical. If there are no extra proteins there are no HIV proteins. If there are no HIV proteins there is no HIV. The particles labelled “HIV” are nothing but cellular microvesicles.168
Bess et al protein profiles of sucrose density gradient banded material from culture supernatants A = uninfected; B and C = “HIV-infected”; Actin and HLA DR = cellular proteins; kDa = molecular weight scale.
In email correspondence Julian Bess told the Perth Group, “We agree that you can come to the conclusion from gel electrophoresis patterns that there are only quantitative differences between HIV and microvesicles [cellular debris]”. If Bess agrees that HIV and cellular material contain the same and same number of proteins, then he must also agree “you can come to the conclusion” there are no HIV proteins and thus no HIV.
Bess also told the Perth Group, "We did not determine the identities of the bands [p6/7, p17 and p24] in this particular gel...these labels were added when one of the reviewers asked for them...He felt it would help orient readers when looking at the figure".
The p6/7, p17 and p24 proteins which Bess and his reviewers assumed are “HIV” proteins, are also present in lesser amounts in the uninfected material. The higher concentrations (darker bands) in the “HIV-infected” material can be explained by differences in the manner in which the cell cultures were obtained and maintained. Since the existence of p24 was proven by Montagnier, and originated in density gradient material in which there were no retrovirus particles, it can only be a cellular protein. And since p6/7, p17 and p24 are derived from a larger, p53 “polyprotein”, p6/7 and p17 are also cellular proteins.
26
- In 1987 Henderson and colleagues169 showed that the “HIV” proteins in the region p30-p32 and p34-p36 region are respectively the alpha and beta chains of the cellular protein HLA- DR. This is confirmed by the annotation in the Bess et al strips.
- Bess labelled a protein with a molecular weight of 41K as actin in both the “purified virus” and uninfected material. Actin is a cellular protein.
- There is no p41 protein labelled “HIV” in either “HIV infected” electrophoresis.
- In 1989 the “HIV” proteins p120 and p160 were shown to be polymers of the p41 protein.170 No proteins of these molecular weights were labelled as “HIV” by Bess and colleagues.
- No p51/p66 (reverse transcriptase) protein was identified in the Bess’ electrophoretic strips.
In summary, the Bess electrophoretic data encompass two striking features. First, density gradient “purified” supernatants obtained by culturing (cells + “virus”) and (cells “– virus”) yield qualitatively identical protein profiles. Any differences are quantitative. Second, only three proteins are labeled “HIV”. These proteins, p6/7, p17 and p24, are present in greater amounts in “infected” material but a higher concentration does not prove they are viral. Bess admitted he had no proof they were viral. This means the “HIV” proteins are cellular proteins. Since there are no “HIV” proteins there can be no HIV antibodies and thus no HIV antibody tests.100, 108-113, 115, 117, 171-176 The role of genes (the genome) is to instruct the synthesis of proteins. If there are no “HIV” proteins, how can there be an “HIV” genome? We will now consider this evidence.
THE HIV GENOME – Fourth major error
It is important to appreciate that the existence of “HIV”, and the HIV theory of AIDS, were accepted before any data were published on nucleic acid sequences claimed to be the HIV genome.
As said earlier, retroviruses have an RNA genome and replicate via a DNA intermediate called the “provirus”. The structure of DNA and RNA is well known. DNA is a double stranded polymer of nucleotides where each nucleotide consists of a nitrogenous base linked to a deoxyribose sugar linked to phosphoric acid. In DNA there are four bases – Guanine, Cytosine, Adenine and Thymine. The two strands in DNA are held together by hydrogen bonding between the bases on opposite strands. Hydrogen bonds are relatively weak and easily disrupted which allows the strands to separate. The nucleotides on adjacent strands pair according to the base pairing rule: G pairs with C and A pairs with T (GC-AT). The sequence of one strand predicts the sequence of the complementary strand. RNA is a single stranded polynucleotide, the sugar being ribose and the nitrogenous base uracil substituted for thymine. An RNA strand can bind to a single strand of DNA following the rule G bonds with C and A with U (GC-AU).
What may not be so well known is nucleic acid hybridisation, a laboratory technique that has been extensively used in HIV research to determine the presence or absence of a given DNA or RNA in vitro and in vivo. The technique is based on complementary base pairing. A radioactively labelled nucleic acid of known sequence (the probe) is added to a mixture of unknown nucleic acids. If DNA or RNA complementary to the probe is present the two strands bind (hybridise). Their union is detected autoradiographically although nowadays radioactive detection has largely been replaced by the use of safer fluorescent or chromogenic markers. If a probe and a DNA hybridise this is often reported as “a positive signal”. Detecting DNA by hybridisation is called Southern blotting after its inventor the biochemist Edwin Mellor Southern. Similar detection of RNA is called Northern blotting.177
Large scale hybridisation studies require large amounts of probe DNAs. This is achieved by cloning. DNA is inserted into the DNA of a microorganism, the latter called a cloning vector, commonly the genome of a bacteriophage (a virus that infects bacteria, also called a phage), often the λ (lambda) phage. After introducing the “recombinant” DNA into a microorganism, often the bacterium Escherichia coli (E. coli), the bacteria are cultured which multiplies the phage with its inserted DNA. The DNA insert is then recovered.
27
Most genomes are too long to work with and thus are first digested (cut into appropriately sized pieces) by enzymes known as restriction enzymes. This results in DNA fragments small enough to be inserted into individual vectors but not so small that individual genes are divided. A collection of different DNA sequences is called a DNA library.178
According to the central dogma of biology, DNA serves as a template for the synthesis of RNA which in turn directs the synthesis of proteins. DNA RNA protein. Many types of RNA are synthesised in the nucleus and much of this is degraded, while others are modified and exported to the cytoplasm. One of the latter is an RNA modified by polyadenylation. Polyadenylation is the addition of an adenine polymer consisting of 100-200 adenine nucleotides to one end of the RNA molecule. The adenine polymer is referred to as the "poly(A) tail" and the RNA as poly(A) RNA. The poly(A) RNA is also called messenger RNA (mRNA) as it directs the synthesis of proteins (translation) within the cytoplasm.179
HIV experts claim it is possible to produce HIV particles by the introduction of unembodied HIV DNA (reverse transcript of the supposed HIV RNA, cDNA) into cell cultures. The experts refer to such DNA as the “infectious molecular clone”. In other words, DNA molecule retroviral particles. This must not be confused with DNA molecular cloning, that is, DNA molecule DNA molecule. To qualify as an infectious molecular clone the introduced DNA must result in the formation of virus particles bearing the same morphological features and biochemical constituents as the virus particles from which the parental RNA (cDNA) was obtained.
As seductive as molecular biology has become, it is important not to equate it with virology. DNA, RNA and proteins are large molecules composed of repeated subunits (polymers). Viruses are infectious particles made of nucleic acid, proteins and other molecules. As Vincent Racaniello teaches his students “A virus is not the same as a virus [nucleic acid] sequence. If you isolate a 200 nucleotide sequence from a specimen that does not mean that the virus is present”.180 In fact this matter was specifically addressed in the Parenzee leave for appeal hearing181 when the Prosecutor presented a paper entitled "Sequence-Based Identification of Microbial Pathogens: a Reconsideration of Koch's Postulates"182 as evidence that genetic methods can be used to prove a virus exists. During cross-examination one of us (EPE) read to the court what the authors stated in their paper: "...with only amplified sequence available, the biological role or even existence of these inferred microorganisms remains unclear" (emphasis ours). Ultimately, the HIV experts, including Gallo, testified that to identify the viral genome the virus particles must be purified. LINK
The HIV genome in vitro The seminal event in identifying an organism’s DNA or RNA is not the cloning and sequencing technology. A police officer wishing to know if DNA collected at a crime scene belongs to a suspect must obtain an oral swab (cells and thence DNA) from that person. And subsequently the law requires an evidentiary chain proving the source of the DNA is the person identified as the suspect. As it is with humans so it is with retroviruses. To define the genome of a retrovirus there must be proof that a retrovirus exists and the nucleic acid comes from that retrovirus. In other words, the scientist must extract and sequence the RNA from purified retroviral particles. Then, the RNA can be none other than the retroviral genome. However, neither Montagnier nor Gallo nor any other HIV researcher defined the “HIV” genome in this manner.
Gallo and his colleagues were the first scientists to undertake experiments claimed to define the HIV genome. His methods and data are reported in three papers published on August 31st, November 8th and December 7th 1984.14-16 Gallo’s evidence offers the most detailed case yet published for the existence of an “HIV” genome and because of its importance, we will first summarise his data before discussing each part in detail.
28
SUMMARY OF THE GENOME EXPERIMENTS
The aim of Gallo’s genome studies was fourfold: identify the HIV genome; prove it fulfilled the retroviral replication cycle; clone the genome; and prove the virus is exogenous. An exogenous retrovirus is one that is acquired from without, that is, external to the cell. As mentioned, “endogenous retrovirus” is a term used to signify the presence of sequences resembling retroviral genomes in human DNA which are not expressed as infectious particles. Such sequences are said to occupy 8% of the human genome.49, 50
- Identify the HIV genome Supernatant from “infected” H9 cultures was banded in a sucrose density gradient and, without any electron microscopic proof it contained retroviral-like particles, the 1.16 g/ml band material was declared to be purified HIV. A poly(A) RNA extracted from the 1.16 g/ml band was then declared the HIV genome. Poly(A) RNA is not specific to retroviruses (see below).
-
Prove HIV satisfies the retroviral replication cycle The retroviral replication cycle consists of four steps:
(a) retroviral particles present in a culture supernatant are taken up by uninfected cells; (b) within the cytoplasm the retroviral RNA is reverse transcribed into its complementary DNA (cDNA). This DNA is unintegrated;
(c) the cDNA is transported to the cell nucleus where it is integrated into the cellular DNA as the “provirus”; (d) the provirus is expressed.
However, any RNA whether incorporated in particles (viral, retroviral or microvesicles) or naked (unembodied), can be taken up by cells and reverse transcribed.183-185 And if retroviral DNA is incorporated into the cellular DNA there is no reason why the same should not occur with any other DNA, complementary or not.100 In other words, the evidence that proved Gallo’s poly(A) RNA, the “HIV genome”, fulfilled the three steps: entry, reverse transcription and integration, does not prove Gallo’s poly(A) RNA is retroviral. - Clone the cDNA of the poly(A) RNA They obtained a number of different cDNA clones but since any DNA, viral or non- viral, can be cloned this does not add support to the claim that their poly(A) RNA is retroviral.
-
Prove the poly(A) RNA is the genome of an exogenous virus This was divided into two parts, in vitro and in vivo. In vitro: A number of uninfected cell lines of different lineage were cultured with supernatant from “infected” H9 cell cultures, that is, with the same supernatant from which their poly(A) RNA (cDNA) was obtained. The cDNA clones were used to probe the DNA of “infected” as well as a variety of non-infected cells. Positive results were obtained only in the case of “infected” cells. However, since the cDNA probes are reverse transcripts of material (RNA) originating in the same supernatant used to “infect” the cells, positive findings would be expected even if both the cDNA probe and the DNA detected in the cells were cellular. This is no different from planting a suspect’s DNA at a crime scene. In vivo: When the same cDNA probes were used to test cells from AIDS patients, that is, cells from individuals who must be infected with HIV, no proof for hybridisation was obtained. In other words, the “HIV genome” was found wherever it was experimentally introduced but not where it should have been introduced by an exogenous retrovirus.
29
GENOME EXPERIMENTS IN DETAIL Experiment 1 (Arya et al)
In this experiment Arya et al describe how they obtained “purified virus” from “infected” H9 cells and then lysed “particles” to extract a poly(A) RNA. The latter was then used as a template to synthesise a complementary DNA. They wrote: "Virus particles were purified from supernatant fluids of HT cells, clone 9 (H9) infected with HTLV-III (HTLV-IIIB) [HIV] by centrifugation through a sucrose density gradient at equilibrium [density gradient banding]”. From the density gradient material (the material Gallo called “purified virus particles”) they obtained a poly(A) RNA. The poly(A) RNA was reverse transcribed into a complementary DNA. In the authors’ words, "The resulting polyadenylate [poly(A)]-containing RNA, was used as template to synthesise 32P- [radioactively] labelled complementary DNA (cDNA) in the presence of oligo(dT) primers. The size of the resultant cDNA ranged from 0.1 to 10 kb (not shown)”.14
Interpretation
The poly(A) RNA is the genome of a retrovirus.
Comments
Arya et al made this claim despite the fact that poly(A) RNA is not retroviral-specific. They published no electron micrographs to prove that the density gradient material (the “purified” virus) consisted of virus particles or any particles of any kind, purified or unpurified. Furthermore, it is not possible to determine how the “HTLV-III” used to infect the H9 cells was obtained in the first place. Different accounts are given in three different papers, although Gallo is a co-author of all of them:
(a) In the Arya et al paper the authors write “HTLV-IIIB [HIV] was originally obtained from pooled supernatants of short-term lymphocyte cultures of AIDS patients" (emphasis ours).
30
- (b) In a paper published in 1985 "The H9/HTLV-IIIB cell line was derived from the human T-cell line HT, following co-culture with T lymphocytes obtained from several AIDS patients, and contains many different HTLV-III forms"186 (emphasis ours).
- (c) In the May 1984 Science papers (where Gallo claimed proof for the existence of HTLV- III) one gets the impression the HT cell line was cultured with concentrated (supernatant) fluids originating from PHA-stimulated T-cell cultures of individual AIDS patients. However, the two year National Institutes of Health Office of Scientific Integrity (OSI) investigation into Gallo’s alleged scientific misconduct and laboratory practices found that the HT cell line was cultured with pooled culture fluids.109, 156, 187, 188 Initially, specimens from three and ultimately ten patients contributed to the mixture.187 The Gallo investigation found this to be "of dubious scientific rigor". One scientist labelled the procedure "really crazy".158 The unnamed scientist speaks for many. No physician in a lifetime of practice would contemplate diagnosing a patient, for example, one with pulmonary tuberculosis or a urinary tract infection, by attempting to isolate a bacterium from a culture of a patient’s sputum or urine mixed with those of nine other patients. It is a particularly pointless experiment in the case of retroviruses because, as retrovirologists have long known, the mere act of co-culturing may lead to de novo appearance of retrovirus-like particles.189, 190 In evidence given to the OSI investigation Popovic said he pooled the supernatant fluids from the ten cultures because none "individually was producing high concentrations of reverse transcriptase".158, 191 It is important to note that a non-specific, background level is inherent in Popovic's method of determining reverse transcriptase activity, and that this level must be exceeded to qualify as the "high concentrations" Gallo and his associates decided were proof that a retrovirus was present. However, since none of Popovic’s ten cultures produced such "high concentrations", none were infected with a retrovirus. One cannot create a retrovirus by manipulating ten sources where no retrovirus is present to begin with.
Experiment 2 (Arya et al)
In this experiment poly(A) RNAs were obtained from (a) “HIV infected” H9 cells; (b) a leukaemic cell line; (c) HTLV-I and -II cell lines; (d) uninfected H9 cells. These RNAs were then probed with 31
the cDNA obtained in the first Ayra et al experiment. Positive hybridisation signals were found only with the poly(A) RNAs obtained from “HIV infected” cells. They detected several “RNA species of about 9.0, 4.2, and 2.0 kb” and concluded, “These bands are similar in size to those corresponding to genomic size messenger RNA (mRNA) and spliced mRNAs of env and pX sequences previously observed in cells infected with HTLV-I, consistent with the relatedness of these viruses".
Interpretation
The poly(A) RNA which bands at 1.16 g/ml band is the genome of an exogenous retrovirus.
Comment
Since the nucleic acids used to obtain the probes and to “infect” the cell lines originated in the same supernatant, one would expect positive hybridisation with the “infected” cells even if the nucleic acids were non-viral (see below). However, if the probes were retroviral one would expect a positive hybridisation with the non-infected H9 cells. This is because (a) as far back as 1983, in a survey of human leukaemias for human retrovirus sequences, Gallo showed that HUT 78, the ancestor of the H9 cell line, "contained HTLV proviral sequences";159 (b) in the Ayra et al paper Gallo and his colleagues have “shown by nucleic acid hybridization that sequences of the genome of HTLV-III are homologous to the structural genes (gag, pol, and env) of both HTLV-I and HTLV-II and to a potential coding region called pX located between the env gene and the long terminal repeating sequence that is unique to the HTLV family of retroviruses”. In fact this is asserted in the title of the Ayra et al paper “Homology of Genome of AIDS-Associated Virus with Genomes of Human T-Cell Leukemia Viruses”.
Experiment 3 (Hahn et al)
Hahn, Gallo and their colleagues replaced the production of cDNA by reverse transcription of poly(A) RNA with molecular [DNA] cloning. “Fresh uninfected H9 cells (8 X 109) were infected with concentrated supernatant from cell line H9/HTLV-III containing 4x1011 particles of HTLV- III...Extrachromosomal [cytoplasmic] DNA was extracted...and assayed for its content of unintegrated viral DNA using HTLV-III cDNA as a probe”.15 (The probe was the HIV cDNA
32
obtained in experiment 1. Unintegrated refers to DNA not inserted into nuclear DNA). “A band of ~ 10 kilobases (kb)” was found to hybridise from which the authors concluded this “DNA represents the linear form of unintegrated HTLV-III [HIV]”. When they digested the unintegrated DNA with restriction enzymes it generated “three predominant bands of 9, 5.5 and 3.5 kb”. They “cloned this DNA into a λ phage library to be screened with viral cDNA”. They obtained three clones, λBH-10, λBH-5 and λBH-8 which they interpreted as “λ BH5 plus λ BH8 constitute one HTLV-III genome and λBH-10 another...However, the viral fragments cloned into λ BH5 and λ BH8 may have been obtained from the same or two different viruses” and are “two variant forms of HTLV-III in the H9/HTLV-III cell line”. They also stated, “Using these clones as probes, we also detected HTLV-III viral sequences in infected cell lines other than H9/HTLV-III that were established from different AIDS patients, and in fresh uncultured lymphoid tissues of AIDS patients19”. (Reference 19 is the third paper on the HIV genome published by Shaw and his colleagues,16 see below).
Interpretation
“These findings suggest that the cloned HTLV-III genomes reported here represent the probable aetiological viral agent of AIDS”.
Comments
1. According to Hahn et al the H9 cells “were infected with concentrated supernatant from cell lines H9/HTLV-III containing 4X1011 particles of HTLV-III”. No mention is made or evidence provided in any of Gallo’s publications showing how the existence, number, morphology or purity of particles were determined.
- In the first paper (experiment 2) the cDNA was used to probe “infected” and non-infected cells. In the Hahn et al paper three clones of unintegrated DNA were used for the same purpose. These differences are methodological and neither paper proves the DNA is that of a retrovirus.
- Even if these data were proof for the existence of a retroviral genome they do not prove the virus is exogenous (see above), much less the cause of AIDS.
Experiment 4 (Shaw et al) The Shaw et al paper16 (Hahn’s reference 19) repeated the methods used in the Hahn et al paper to obtain what they claimed to be two DNA clones of the integrated viral genome. That is, DNA inserted in the nuclear DNA as “the provirus”. These clones were called λHXB-2 and λHXB-3 and since 1984 commonly used in HIV genomic studies as well as for the diagnosis and monitoring of HIV infection. “To determine whether the HTLV-III genome contains sequences homologous to normal human DNA”, λHXB-2 was used to probe HTLV-III infected and uninfected cellular DNA. “Under standard conditions of hybridization...this probe hybridized to DNA from H9/HTLV-III cells as well as other HTLV-III-infected cells, but not to DNA from uninfected H9 cells, uninfected HT cells (the parent cell line from which H9 was cloned), or normal human tissues (data not shown)”. In fact, Shaw and his colleagues were the first to use HXB-2 for diagnostic purposes. That is, to detect the purported HIV genome in AIDS patients (see below).
Interpretation
“This finding is in agreement with the results of other experiments [reported in Hahn et al paper] in which the unintegrated (replicative intermediate) form of HTLV-III was used as probe and demonstrates that HTLV-III, like HTLV-I and HTLV-II, is an exogenous retrovirus lacking nucleic acid sequences derived from human DNA”.
Comments
The method and findings in this experiment are no different from those of their previous experiments. This being the case, the results of this experiment do not prove that the virus is exogenous.
33
GENERAL COMMENTS ON THE IN VITRO GENOMIC EXPERIMENTS Like Montagnier, Gallo failed to use valid controls. This omission is especially significant given that retroviral genomes are present “in all of us”.49
Poly(A) RNA and the “HIV genome”
-
All experiments described above are based on RNA obtained from material referred to as “purified virus”. Yet, as repeated ad nauseam, the “purified virus” was none other than the 1.16 g/ml sucrose density gradient band. Furthermore:
(a) at no time was there proof the 1.16 g/ml band material used in these experiments contained retroviral particles, or particles of any kind, pure or impure;
(b) like Gluschankof and his colleagues, until 1997 every HIV researcher including Gallo and his colleagues, must have known that "the purity of the virus preparation [has not] been verified”;160
(c) it had long been known that the 1.16 g/ml band may contain cellular RNA either embodied (a constituent of cellular microvesicles and debris) or unembodied (free). In 1975 the distinguished retrovirologist John Bader wrote, "Unfortunately, cellular components also can be found banding at the density of the [retro]virus, especially membraneous vesicles which may enclose other cellular constituents, including nucleic acids...[RNA and DNA] No published technique has over-come these encumbrances to virus purification. In lieu of documentation to the contrary [which is why electron microscopy is crucial], one must assume that all preparations of virions contain contaminating cellular elements".77 Two decades later Bess et al confirmed Bader’s reservations – “Microvesicles [which band at the retroviral density of 1.16 g/ml] were found to contain various proteins...and a substantial amount of RNA and DNA” including messenger RNA, that is, poly(A) RNA.162 - Justification for defining poly(A) RNAs as retroviral HIV RNA was based on work done in 1972 by researchers (Gallo and colleagues included) one year after the discovery of poly(A) RNA. Because retroviral RNAs were shown to be poly(A) in type it was concluded that poly(A) RNA is a diagnostic property of retroviruses.192 However, like reverse transcriptase and the earlier “retrovirus-associated” enzyme ATPase,193 poly(A) RNA is not retroviral-specific. In 1980 such evidence was available to all HIV researchers. Indeed, in an extensive review article "A history of poly A sequences: from formation to factors to function", Mary Edmonds, who discovered the enzyme that catalyses the addition of the poly(A) tails to RNA, wrote, "poly-A sequences were found in both messenger RNA (mRNA) and their nuclear [cell nucleus] precursors...poly(a) sequences provided the basis for a long-sought route for mRNA purification...and generating the cDNAs and the probes derived from them on which so many studies of gene expression continue to depend".194
- If the sucrose density band from which the poly(A) RNA originated had contained "purified virus” particles then the band should have consisted of poly(A) RNA and no other RNA or DNA. There should have been no need to target (and extract) poly(A) RNA.
-
In all their papers Gallo and his colleagues repeatedly state that the purified virus particles were obtained by banding in "sucrose density gradients". However, during the Parenzee court case of 2006/2007 Gallo was asked if in 1983 Montagnier had purified HIV. He replied: "He did a 116 cross gradient [1.16 g/ml sucrose density gradient] in that paper, yes. I don't know if he said it was purified. If you do that you don't have
34
much virus".195 Yet Gallo and his associates did exactly that and by his own reasoning he also “[didn’t] have much virus”.196
5. In February 2003 a paper197 published in the British Medical Journal sparked an intensive, 26 month long, 842 postings on-line debate that was terminated by the editor in April 2005. LINK During this debate we had several exchanges with Brian Foley, custodian of the Los Alamos HIV Database who ultimately agreed that:
- (a) Gallo's poly(A) RNA is at present considered to be the HIV genome;
- (b) poly(A) RNA is not specific to retroviruses;
- (c) the poly(A) RNA originated from the 1.16 g/ml sucrose density band;
-
(d) there was no proof the 1.16 g/ml density band contained purified retroviral
particles, or any retroviral particles, pure or impure.
However, Foley still insisted that Gallo's poly(A) RNA is the HIV genome. He based this assertion on his claimed existence of the HIV infectious molecular clone. That is, he claimed there are published data showing that the introduction of that RNA or its complementary DNA into a cell culture results in the production of retrovirus particles bearing the same morphological features and constituents as “the parental virus” particles. When we asked for proof for the existence of such an HIV infectious molecular clone he responded with a long list of papers. Although the titles of these papers included the phrase "infectious molecular clone" no such evidence could be found in any of them. All they had was proof for DNA cloning, that is, the production of DNA from DNA.
We asked Foley the following question:
Is it true that by infectious molecular clone of a virus is meant the introduction of the viral genome (molecular clone) into suitable cells leading to the appearance of virus particles identical (regarding both appearance and composition) to the ones from which the genome was obtained? Yes or No?
Foley agreed there is a difference between cloning a DNA fragment and an infectious molecular clone. He even gave us his own definition of the latter:
Yes, as I have said twice or more times before already. The clone must produce virus particles that are identical by serology, morphology, protein sequences, RFLP [restriction fragment length polymorphism, a largely obsolete method of DNA profiling], Southern blotting, etc. to the parental virus, and the particles must also be infectious. If a cloned viral genome does not meet these criteria, it is not an INFECTIOUS molecular clone of the virus, be it HIV-1 or any other virus. Morphology by electron microscopy is the least important of those criteria, because as I have stated many times before, all lentiviruses look alike by electron microscopy. Also, it is not always possible to detect the difference between infectious and non-infectious virus particles by electron microscopy.
In fact, it is not possible to prove the existence of an infectious molecular clone using Foley’s criteria. To prove particles are identical would require purification not once, but twice. The first time to obtain and characterise the parental retroviral RNA, and the second to prove the RNA from the “infectious molecular clone” is identical. There has never once been evidence for purification of any “HIV” particles.
We then asked Foley for proof for the existence of the HIV infectious molecular clone according to his definition:
Would Brian Foley please give us a study and a few confirmatory studies where the existence of an “infectious molecular clone” of “HIV” has been proven.
35
If he failed to do so then obviously he would have no choice but to accept that the existence of an HIV genome and thus a retrovirus HIV remains unproven. When we realised Foley was unable to produce such proof we wrote:
So we repeat our request: Would Brian Foley please give us a summary of the evidence (not just the title) of a study as well as the evidence from a few confirmatory studies where the existence of an “infectious molecular clone” (as defined by Brian Foley) of “HIV-1” has been proven. If Brian Foley fails to respond with his summaries and references then we must conclude his whole argument for the existence of “HIV-1”, based upon the existence of the “HIV-1 infectious molecular clone”, collapses.
Instead of giving us the proof we requested according to his own criteria, Brian Foley, Simon Wain- Hobson and John Moore put pressure on Richard Smith, editor of the British Medical Journal, to stop the debate. Inexplicably, the pressure came not via the British Medical Journal but through Nature.93 Smith’s response included, "As editor of the BMJ, however I find it disturbing to see scientists arguing for restriction on free speech. Surely open communication and argument is a fundamental value of science...We should never forget Galileo being put before the inquisition. It would be even worse if we allowed scientific orthodoxy to become the inquisition".198 Shortly after Richard Smith’s resignation as Editor in Chief, the BMJ’s letters editor, Sharon Davies,199 terminated the debate. By their contribution to terminating the debate Foley, Wain-Hobson and Moore ignored the advice of the father of modern retrovirology, the Nobel laureate the late Howard Temin: “When an experiment is challenged no matter who it is challenged by, it’s your responsibility to check. That is an ironclad rule of science, that when you publish something you are responsible for it...even the most senior professor...if challenged by the lowliest technician or graduate student, is required to treat them seriously and consider their opinions...It is one of the most fundamental aspects of science”200 (emphasis in original). In 2010 the BMJ published a paper whose authors include Smith’s successor Fiona Godlee. Ironically the paper concluded "Authors are reluctant to respond to criticisms of their work...Editors should ensure that authors take relevant criticism seriously and respond adequately to it”.201
The in vitro evidence for the exogenous nature of the HIV genome
- Since nucleic acids present in the same supernatant were used to obtain the probes (cDNA and its clones) and to infect H9 and other cells, one would expect positive hybridisation with “infected” cells irrespective of the origin of the nucleic acids, viral or non-viral.
- Gallo and his colleagues claim cells were infected using supernatant “containing 4X1011 particles of HTLV-III”. No mention is made of the volume and more importantly, how they determined there were “4X1011 particles” or indeed any particles. Given that only months earlier Gallo’s electron microscopist Mathew Gonda had problems finding particles in the supernatant of infected cells, the absence of electron microscopic evidence is pivotal.156
The facts are:
- Gallo never had any proof that the “HTLV-III RNA” and thus the cDNA and its clones originated in particles with the morphology of retrovirus particles much less infectious retroviral particles.
- Poly(A) RNA is not specific to retroviruses.
- The 1.16 g/ml band obtained by density gradient centrifugation of non-infected cell culture supernatants contains poly(A) RNA , as shown by Bess and his colleagues.162
-
As mentioned, irrespective of its origin, any RNA or DNA present in a supernatant can be taken up by cells and reverse transcribed.183-185 If retroviral DNA can be incorporated into
36
the cellular DNA there is no reason why the same should not happen with any other DNA.100
- Reverse transcription is not retrovirus specific.
- At present it is still generally accepted that any time one finds a novel DNA in a cell, for example, in a T-lymphocyte, the interpretation is that the DNA has been introduced from outside. However, according to Barbara McClintock, the genome can be restructured and not only by transposition (the removal and relocation of a DNA sequence into another place in the genome). In her 1983 Nobel lecture she said, “rapid reorganisation of genomes may underline some species formation. Our present knowledge would suggest that these reorganizations originate from some “shock” that forced the genome to restructure itself in order to overcome a threat to its survival...Major genomic restructuring most certainly accompanied formation of new species”. The “genomic shock” which leads to the origin of new species may be “either produced by accidents occurring within the cell itself, or imposed from without such as virus infections, species crosses, poisons of various sorts, or even altered surroundings such as those imposed by tissue culture”202 (emphasis ours).
- Evidence published since then by those in the forefront of genetic research vindicates McClintock’s views. This is especially the case with the evidence published by scientists involved in the human genome project which was undertaken to map the human genes and usher in genetically based, personalised medicine; notably the evidence gathered by the 300 scientists from 10 countries participating in the Encyclopaedia of DNA Elements (ENCODE) project.203, 204 The aim of ENCODE is “to identify all regions of transcription, transcription factor association, chromatin structure and histone modification in the human genome sequence”.203 According to the ENCODE researchers it is not possible to say where transcription of DNA begins and ends, that is, it is not possible to say what a gene is.205 “The picture these studies paint is one of mind-boggling complexity. Instead of discrete genes dutifully mass-producing identical RNA transcripts, a teeming mass of transcription converts many segments of the genome into multiple RNA ribbons of differing lengths. These ribbons can be generated from both strands of DNA, rather than from just one as was conventionally thought...We’ve come to the realisation that the genome is full of overlapping transcripts...the simple view of the genome as having a defined set of isolated loci transcribed independently does not seem to be accurate. Protein-coding sequences have no clear beginning or end”.206 In fact as Denis Noble points out, “Some biological scientists have even given up using the word ‘gene’, except in inverted commas”.207 More than thirty years ago, one of us (EPE) suggested that the biological dogma is flawed and there is feedback between the three polymers, DNA, RNA and proteins, regulated by the cellular redox and its oscillation, in particular, a myosin/actin redox couple.208
-
At present it is believed that any time one finds a particular sequence of RNA in a cell, for example, a T-lymphocyte, unless the RNA has been introduced from outside, all cells, regardless of their physiological state or stresses, will contain a corresponding stretch of DNA. In other words, the DNA (genes) in a cell is invariant and all RNA molecules in the cell are accompanied by a matching length of DNA. This is not the case. In the 1980s RNA editing was discovered. This is “broadly defined as a process that changes the nucleotide sequences of an RNA molecule from that of the DNA template encoding it”. In the process a non-functional transcript can be re-tailored, producing a translatable mRNA, or modify an already functioning mRNA so that it generates a protein of altered amino acid sequences. Sometimes editing is so extensive that the majority of sequences in an mRNA are not genomically encoded but are generated post-transcriptionally producing the “paradoxical situation of a transcript that lacks sufficient complementarity to hybridize to its own gene!”.209-211 Evidence published more recently shows that RNA editing has a
37
significant role in many cellular and biological phenomena and can be induced by chemical stress.212-215
- None other than Montagnier agrees that novel RNAs can arise without the agency of exogenous infectious agents. In a written testimony dated February 2nd 2000 to the US House of Representatives Committee on Government Reform, Subcommittee on National Security, Veterans Affairs and International Relations, in support of the work of his colleague, Howard B Urnovitz, (Montagnier is on the scientific advisory board of a publicly traded biomedical company whose director is Urnovitz), Montagnier wrote: "I have reviewed Dr Urnovitz's published research and the testimony prepared for presentation to this Committee and strongly advise that future research on Gulf War Syndrome should include the study of the detected genetic material". Urnovitz and his colleagues presented evidence of the existence, in Persian Gulf War veterans, of "novel", "nonviral" RNAs, "possibly induced by exposure to environmental genotoxins". They concluded: "The patterns of the occurrence of RPAs [polyribonucleotides] in the sera of GWVs [Gulf War Veterans] and healthy controls are sufficiently distinct to suggest possible future diagnostic applications...Our studies of patients with active multiple myeloma suggest that patients with individual chronic multifactorial diseases may have unique RPAs in their sera. Validated tests for such putative surrogate markers may aid in the diagnosis of such diseases or in the evaluation of responses to therapeutic modalities".216
- Gallo obtained his poly(A) RNA (the HIV genome) from cells kept in culture for long periods and exposed to strong oxidising agents (“shocks”) including PHA. PHA is an oxidant ubiquitous in HIV research and causes “Both the levels of RNA proteins synthesis [to] increase drastically” and the appearance of novel mRNA (poly(A) RNA).217, 218 Not surprisingly, in these cultures one will expect to find “novel” mRNAs (“HIV” RNA) which cannot be found in cultures not exposed to similar “stress”. Since the PCR hybridisation probes originate from these cultures and since AIDS patients and those at risk are exposed to similar oxidative stress, one would predict these patients will have a positive PCR test (“viral load”26) which the HIV experts attribute to “HIV” infection. Since ARV drugs are also oxidising agents65, 219-221 they can be expected to cause further RNA editing and histone and chromatin modifications. Given that the PCR probes originate from cultures which were not exposed to the extra “stress” of ARVs, one would expect the production of RNAs that do not complement with the primers employed in the amplification step in the “HIV” RNA assay. By this means the “viral load” will decrease or even become non-detectable when patients are treated with antiretroviral compounds.222
- Whether McClintock’s “shock” induced “rapid reorganisation of genomes”, or “many segments” of DNA transcription, or RNA editing or combinations involving all three, the discovery of a new RNA transcript does not prove the existence of a continuous, complementary DNA sequence within the nuclear DNA; or that the RNA was introduced from outside the cell. In fact, the one thing about which the ENCODE authors were certain was that “Chromatin accessibility and histone modification patterns are highly predictive of both the presence and activity of transcription start sites” and “DNA replication timing is correlated with chromatin structure”. Since histone regulation and chromatin structure are redox dependent,208, 223, 224 and HIV-positive and AIDS patients have perturbations in cellular redox (which occurs early and is “massive” according to Montagnier225), ENCODE affords a non-virological explanation for the generation of not just the original RNA 6 transcript but also the awkward fact that “an asymptomatic patient can harbour at least 10 genetically distinct variants of HIV, and for an AIDS patient the figure is more than 108”.226,
227 And all claimed to be the genome of a “unique” retrovirus (see below).
38
THE HIV GENOME IN VIVO The acid test of HIV and the HIV theory is proof that the HIV genome is present in the T4 cells of all AIDS patients. All the in vivo genomic data were published by Gallo and his colleagues. Montagnier's group did not mention any attempt to find and characterise the HIV genome in AIDS patients.
According to Gallo hybridisation is so sensitive it can detect a DNA in one in a million cells. Hence, since:
- All individuals with HIV or AIDS should be infected with HIV (and hence have detectable HIV proviral DNA); 87 88
- According to the HIV experts David Ho and Xiping Wei, HIV-positive individuals have massive HIV infection from inception and “The estimated average total HIV-1 production was 10.3 x 10(9) [109] virions per day”,228
Southern blot hybridisation should be more than adequate to detect it.
But in vivo detection of HIV DNA is problematic. Despite the Hahn et al claim that Shaw et al found HIV sequences “in fresh uncultured lymphoid tissues of AIDS patients”, in the latter paper the best Gallo and his colleagues could say was: "...as shown herein, HTLV-III [HIV] DNA is usually not detected by standard Southern Blotting hybridization...when it is, the bands [signals] are often faint...the observation that HTLV-III sequences are found rarely, if at all, in peripheral blood mononuclear cells, bone marrow, and spleen [T4 cells or anatomical sites where T4 cells are located] provides the first direct evidence that these tissues are not heavily or widely infected with HTLV-III in either AIDS or ARC [AIDS related complex = a non-specific AIDS prodrome]".16 The finding of "faint", "low signal" hybridisation was interpreted as “This must mean that only a minor population of cells is infected with HTLV-III at any one time” and “Theoretically, this low signal intensity could also be explained by the presence of a virus distantly homologous to HTLV-III in these cells”, HTLV-I or HTLV-II. In other words, Gallo was not able to prove the existence of the HIV genome in AIDS patients.
While Gallo was unable to prove the existence of the HIV genome in AIDS patients, others reported “HIV” DNA hybridisation signals in situations where the absence of an HIV genome is not disputed. For example:
- Although it is no longer accepted that “HIV” is transmitted by insects, in 1986 using Southern Blot Hybridisation several of Montagnier’s colleagues at the Pasteur Institute found what they interpreted as HIV DNA sequences in tsetse flies, black beetles and ant lions in Zaire and the Central African Republic.229
- In 1984 Weiss and his colleagues reported a retrovirus in two young, HIV-negative adults who had the disease common variable hypogammaglobulinaemia. The retrovirus “was clearly related to HTLV-III/LAV [HIV]”. Their evidence included Southern blot hybridisation using Gallo’s HXB-2 (HIV) as the probe.230
- DNA extracted from thyroid glands from five HIV-negative patients with Grave’s disease (thyrotoxicosis) hybridises with “the entire gag p24 coding region of the HIV-1 genome”.231
- Horwitz et al, “describe the first report of the presence of nucleotide sequences related to HIV-1 in human, chimpanzee and Rhesus monkey DNAs from normal uninfected individuals”.232
The absence of an HIV genome in AIDS patients was a major problem for the HIV theory and the HIV experts. However, both were soon to be rescued by the development and rapid commercialisation of an ultimately Nobel prize winning technique capable of rapidly producing large quantities of DNA – a technique invented by Kary Mullis called the polymerase chain
reaction233 (PCR). As Mullis himself points out, there are many problems associated with the use 39
of the PCR for HIV testing including the following: "PCR detects a very small segment of the nucleic acid which is part of the virus itself...(Two to three hundred nucleotides is usually chosen out of the several thousand [~10K] in the total retrovirus)...There are many sequence variations among the sequences called HIV. The specific fragment detected is determined by the somewhat arbitrary choice of DNA primers used which become the ends of the amplified fragment. They have to be in the sequence for it to be amplified in the first place, but they can be rather a small part of the total sequence. Any one of them can get you classed as what they consider HIV- positive. And due to the tiny amounts of nucleic acid detectable after many cycles of PCR amplification (after 30 cycles one copy will get you about a billion copies) the test is super- sensitive”.
Despite these cautionary words from the inventor of the technique, finding “Two to three hundred nucleotides...part of the virus...chosen out of the several thousand in the total retrovirus”, became the de facto whole virus genome. The product obtained by the PCR is a DNA but to prove the identity of that DNA, that is, the order of its nucleotides, it must be sequenced. Since the sequencing step is not routine, most often there is no proof that the amplified sequence is “HIV” DNA. It could be a DNA with the same or similar ends as the primers. Mullis wrote, “If a sequence amplifies with primers designed to HIV, it is HIV by definition". But “primers designed to HIV” can amplify non-HIV sequences. For example, in 1989 Andy Shih reported, “HIV unique primers corresponding to the highly conserved reverse transcriptase region function well in the PCR amplification of HeLa [cell] DNA”.234 (HeLa is an immortal cell line obtained in 1951 from a woman with cervical cancer235). Plasma HIV RNA (“viral load”) tests give positive results in non-infected individuals. Manufacturers warn "since their specificity is not well known, these tests must not be used for diagnostic purposes".236 Roche, for example, state that “The Amplicor HIV-1 [RNA] Monitor test is not intended to be used as a screening test for HIV-1 or as a diagnostic test to confirm the presence of HIV-1 infection” (Roche Diagnostic Systems, 06/96, 13-08088-001. Packet Insert). This is corroborated by researchers from the Massachusetts School of Medicine, who state “Plasma viral [RNA] load tests were neither developed nor evaluated for the diagnosis of HIV infection...Their performance in patients who are not infected with HIV is unknown” and their use leads to “Misdiagnosis of HIV infection”.237
NO PROOF HIV RNA/DNA IS UNIQUE
Neither PCR nor Southern blot hybridisation are relevant to “HIV” and the HIV theory of AIDS if there is no proof that HIV RNA/DNA is unique. The internationally renowned Australian HIV/AIDS expert David Cooper stated in evidence to the Parenzee hearing: “Once the virus is purified, it’s then genetically sequenced and those sequences are unique just like every organism on the planet has unique sequences and markers”.238 However, since there is no proof that the virus has ever been purified, there is also no proof that sequences assumed to belong to “HIV” in fact do so.
No two “HIV” DNAs have the same sequence. The variation among sequences is accepted to reach as high as 30-40% (Brian Foley, personal communication and Peter McDonald’s testimony at the Parenzee hearing239). Even 50% variation has been accepted by most researchers, without their questioning whether they are really working with a “unique” viral entity. These variations contrast with the 95-98.5 per cent genetic identity between chimpanzees and humans.240
When renaming LAV (lymphadenopathy-associated virus), HTLV-III (human T-cell lymphotropic virus-III) and other isolates in 1986 as HIV, John Coffin and 11 of his retrovirologist colleagues including Howard Temin, Robin Weiss and Montagnier wrote: "Any future isolates of human retroviruses with clear but limited relationship to isolates of HIV (for example, more than 20 per cent but less than 50 per cent nucleic acid identity) should not be called HIV unless there are compelling biological and structural similarities to existing members of the group".17 How can such dissimilar variants have similar and consistent biological properties, including immunogenicity and pathogenicity?
One cannot find two identical genomes even in the same patient. As mentioned, researchers from
the Pasteur Institute state, “an asymptomatic patient can harbour at least 106 genetically distinct 40
variants of HIV, and for an AIDS patient the figure is more than 108”.226, 227 In one and the same patient the genomic data in monocytes differs from that in T-lymphocytes.241 There are “striking differences” between the proviral DNA and cDNA in one and the same PBMC sample “which could not be explained by either an artefact of reverse transcriptase efficiency or template selection bias”.242 The genetic data obtained in vitro do not correlate with the data obtained in vivo, and “the task of defining HIV infection in molecular terms will be difficult”.243 According to Stephanie Gras, Senior Research Fellow, Department of Biochemistry and Molecular Biology, Monash University, Australia, “HIV can change 100,000 times faster than the [in]flu[enza virus], and we need a new vaccine for the flu every single year”.244 How can such genetic variation be considered the genome of a unique virus? Yet, for over thirty years the same antigens [“gene products”] have been used in the antibody test kits and the same primers and probes in the genomic tests (PCR).
For any test the issue of utmost significance for patients and physicians is that of specificity. Even if HIV were to exist it is vital to obtain proof that the DNA detected in a cell culture or the RNA in the plasma (“viral load”) test is not caused by something other than HIV. PCR is extremely sensitive, that is, given the presence of a given DNA it will almost invariably detect it. What is of paramount importance is whether the test is specific. This means that not only does the test always detect a given DNA, it never detects another DNA. However, the specificity of the HIV PCR has never been determined using HIV isolation/purification as a gold standard. In other words, while the PCR is undoubtedly an outstanding means for detecting tiny quantities of DNA, what or whose DNA is it? PCR is a laboratory test and like all tests in clinical medicine should not be used to diagnose or treat patients before its specificity has been determined against a gold standard.107 The only gold standard for HIV is HIV itself, that is, HIV isolation/purification.
In 1996 Douglas Owens and his colleagues at several US institutions addressed the parameters of the HIV PCR test. They wrote, "To evaluate the sensitivity and specificity of PCR, investigators must ascertain whether study participants are infected with HIV. Typically, a new test is compared with a superior reference (or gold standard) test...The lack of an appropriate reference test substantially complicates evaluation".245 The fact is, there is a “superior reference (or gold standard) test” for HIV PCR: HIV itself, that is, HIV isolation/purification. If Owens admits to there being no gold standard for the PCR then it follows there is no virus. This being the case it is impossible to claim the test parameters for the HIV PCR have been determined, and therefore this test should not have been introduced into clinical practice.
Even if the PCR were evaluated using HIV isolation as the “superior reference”, its specificity still could not be determined for the simple reason that the PCR is not standardised. As Owens et al reported, “The criteria for determining when PCR gave positive results varied among the studies”. That is, a person could have a positive PCR in laboratory A but not in laboratory B. Even when the lack of standardisation is ignored and totally unsuitable gold standards are used to try to assess specificity, such as the antibody test,110, 112, 113, 246 Owens reported “specificities range from 40% to 100%”. This means that in some studies 60% of persons not infected with HIV according to the “superior reference” had a positive PCR.247 The authors concluded, “Our investigation produced two main findings. First, the false-positive [negative antibody test/positive PCR] and false-negative [positive antibody test/negative PCR] rates of PCR that we determined are too high to warrant a broader role for PCR in either routine screening or in the confirmation of diagnosis of HIV infection. This conclusion is true even for the results reported from more recent, high-quality studies that used commercially available, standardised PCR assays...We did not find evidence that the performance of PCR improved over time”.245, 248
Christine Defer from the Centre Régional de Transfusion Sanguine reported similar findings from seven French laboratories with extensive experience in HIV PCR technology: “False-positive and false-negative results were observed in all laboratories (concordance with serology [antibody tests] ranged from 40 to 100%)...the number of positive PCR results did not differ significantly between high- and low-risk seronegatives”.249 As far as the “superior reference (or gold standard test)” is
41
concerned, Owens and Defer and all HIV experts use the antibody tests245, 250 despite the fact not one antibody test has been verified against HIV isolation/purification.251
In the 2012 edition of Harrison’s Internal Medicine Fauci asserted “a positive EIA [enzyme immunoassay] with a confirmatory Western blot remains the “gold standard” for a diagnosis of HIV infection”.143 In June 2014 the CDC withdrew the Western blot from the HIV antibody testing algorithm on the basis of “improved immunoassays” and “an HIV-1 NAT [nucleic acid test] (PCR)”.252, 253 In the updated algorithm NAT is the final arbiter of HIV infection, that is, as of June 2014 the PCR dictates the “true infection status” of individuals who have indeterminate antibody tests. In other words, whereas formerly the confirmatory Western blot was the “gold standard” for the diagnosis of HIV infection and NAT (PCR) was not used diagnostically, since 2014 the PCR has become the “gold standard”. Yet at no stage in the history of AIDS have the antibody tests or the PCR been verified against HIV isolation/purification, that is, against the putative virus for which the tests are employed.
ADDITIONAL EVIDENCE THAT “HIV” DNA IS NOT UNIQUE
In September 1998 US researcher Eva Rakowicz-Szulczynska reported the presence of HIV-1 DNA sequences in patients with breast, genital tract and prostate cancers. Using HIV-1 gp41- derived primers she PCR amplified DNAs from tumour tissue of 40 patients with breast cancer. All samples were positive. DNA fragments amplified from seven blindly selected breast cancer samples were sequenced and found to be 141-143 base pairs in length with 90-95% identity to the HIV-1 gene for gp41.254, 255 She obtained the same results with the same primers in patients with prostate cancer. In fact, four years earlier Rakowicz-Szulczynska’s published data showing that a monoclonal antibody directed against the purported HIV-1 envelope gp120 protein reacts with p24, p41, p120 and p160 proteins found in breast and gynaecologic cancers but not in other cancer types or normal tissues.256 Despite her DNA data and finding HIV-1 antibody reactivity with four proteins of the same molecular weights as HIV-1 proteins, Rakowicz-Szulczynska was reluctant to conclude her patients HIV-1 infected. Instead, she opted for infection with a virus showing “epitopic [antigenically similar] and genetic homology to HIV-1”257 and concluded that her findings “strongly suggest that a retrovirus related to HIV-1 may be associated with cancers of the reproductive system”.258
Rakowicz-Szulczynska’s sequences are deposited at the National Center for Biotechnology Information nucleotide database. According to Brian Foley, custodian of the Los Alamos HIV database, these DNA sequences are “clearly subtype B [HIV-1], which is typical for North America” (Brian Foley, personal communication). Foley initially viewed Rakowicz-Szulczynska’s findings as contamination. However, he later agreed with the Perth Group that contamination does not explain why none of the control, non-cancerous tissues adjacent to the tumours were positive for HIV-1 DNA. Rakowicz-Szulczynska’s sequence data can be confirmed HERE (Despite what HIV-1 sequences may signify in regard to the HIV theory of AIDS, given the prevalences and prognoses of breast and gynaecological cancers, Rakowicz-Szulczynska’s findings are potentially a useful tool for the early diagnosis of such neoplasms,259 especially as a liquid biopsy).
In March 2014 Spanish engineer and bioinformatics260-262 graduate Miguel Romero Fernández- Bravo from the Open University of Catalonia, Spain, published data that show the presence of HIV- 1 DNA in a wide variety of non-HIV sources.263, 264 More recently Romero posted additional taxa whose DNAs also bear high identities to HIV-1 DNA. These include human exons, the human prokr2 gene, mosquito and metagenomes (environmental samples) from soil, freshwater, sludge, salterns (a place where salt is obtained from pools of evaporated sea water) and corals. Data HERE Although raising the possibility of another mechanism, Romero interpreted his findings as contamination. Contamination of DNA databases is a risk265 which is why the National Center for Biotechnology Information (NCBI) filters every submitted sequence266 to exclude, as one of many examples, the common bacterium E. coli. However, the NCBI does not publish data on the prevalence of contamination of HIV sequences, nor filter for HIV sequences.
42
Romero subsequently reported HIV-1 DNA in the genome of James Watson267, co-discoverer of the structure of DNA268 as well as in malignant melanoma and several other malignancies: kidney, colon, bone, leukaemia and lymphoma.269, 270 Data HERE These include sequences exceeding 100 nucleotides with 100 percent identities to HIV-1. Again it might be argued that all instances of HIV-1 sequences detected outside HIV-infected individuals are caused by contamination or the use of lentiviral cloning vectors. However, the ease with which one can find HIV-1 DNA in human neoplastic tissues means either contamination is unprecedented or that the nucleic acid sequences are not HIV-specific. Despite the enormous amount of evidence of non-specificity, if a section of HIV DNA is found, for example, in a homosexual man or the newborn of an HIV-positive mother, it is always considered evidence of HIV infection. (It is the only procedure used to diagnose HIV infection in neonates and infants271).
In their 1984 genome experiments Gallo and his colleagues showed that the HIV genome did not hybridise with DNA from uninfected cells. From this they concluded, “HTLV-III [HIV-1] is an exogenous human retrovirus...[and] lacks nucleic acid sequences derived from normal human DNA”.16 However as mentioned, in 1986 and 1989 there were reports showing HIV sequences are not lacking in DNA from humans with certain diseases, including common variable hypogammaglobulinaemia and thyrotoxicosis230, 231 Then, Rakowicz-Szulczynska (2000) and Romero (2015) documented HIV-1 nucleic acid sequences in malignancies as described above. Cancer and AIDS share a common property – cellular oxidation.223, 272-281 Cellular redox underlies cellular structure and function223, 282-284 and from oxidative “shocks...imposed from without such as...poisons of various sorts, or even altered surroundings such as those imposed by tissue culture...the genome can be restructured”.202 Consequently, the detection of one or other “HIV genetic sequences” may reflect the common property of AIDS and cancers and therefore find empirical use as a laboratory test for certain morbid and pre-morbid states. For example, as a liquid biopsy in the diagnosis and treatment of cancers.285-288 (This may well have a serological parallel in the findings mentioned above of St. Louis et al146 who reported a “very high [HIV-1] seroprevalence at some sentinel Hospitals” (26 hospitals in 21 cities) testing 89,547 patients in whom over one hundred diseases or conditions commonly and remotely associated with AIDS were excluded145).
SEX AND HIV – Fifth major error
According to the HIV experts, sexual intercourse is the principal route of HIV transmission. However, there is no microbiological proof based on the isolation of HIV from genital secretions of index cases followed by sexual contact tracing and testing.115 The evidence claimed to prove sexual transmission consists of epidemiological studies which document the relationships between different types of sexual activity and the presence or development of HIV infection, as assumed to have been indicated by a positive antibody test. Even if one accepts that HIV antibody testing is highly specific, the epidemiological data obtained are inconsistent with the behaviour of a sexually transmitted disease or agent.
The sine qua non of sexually transmitted diseases is their bidirectional transmission, that is, from the active (insertive) to the passive (receptive) partner and vice versa. The active partner is the penis inserting, semen donating homosexual or heterosexual male. The passive partner is the penis accepting, semen accepting homosexual or heterosexual female. Proof that a disease is sexually transmitted requires demonstration of a chain of transmission and acquisition from active partner Apassive partner Bactive partner C and so on.
The first to study the relationship between HIV (a positive antibody test) and sexual activity in homosexual men was Gallo and his associates. In a 1984 cross-sectional study they reported that "of eight different sex acts, seropositivity [a positive antibody test] correlated only with receptive anal intercourse".289 In their updated study published in 1986 Gallo wrote: "Data from this and previous studies have shown that receptive rectal intercourse...is an important risk factor for HTLV-III [HIV] infection...We found no evidence that other forms of sexual activity contributed to the risk".290 Unquestionably, the largest, longest, best designed and executed study in
43
homosexual men is the Multicenter AIDS Cohort study (MACS291). Amongst their 1300 scientific publications (which include prospective studies) there are many that confirm "receptive anal intercourse was the only sexual practice shown to be independently associated with an increased risk of seroconversion to HIV in this study".292, 293
By 1994 many epidemiological studies including prospective studies had been conducted in homosexual men. Reviewing more than 20 such studies Caceres and van Griensven concluded: "the cited reports yield convincing evidence that unprotected anogenital receptive intercourse poses the highest risk for the sexual acquisition of HIV-1 infection...there is mounting epidemiological evidence for a small risk attached to orogenital receptive sex, biologic plausibility, credible case reports and some studies show a modest risk, detectable only with powerful designs;...no or no consistent risk of the acquisition of HIV-1 infection has been reported regarding insertive intercourse and oro-anal sex".294 In heterosexual studies the evidence is identical. In other words, the only sexual risk factor for the acquisition of a positive antibody test is passive anal intercourse.295-297
Since HIV infection is synonymous with a positive antibody test and since a positive antibody is acquired only by the passive sexual partner, HIV cannot be a sexually transmitted infectious agent. This means “HIV” is either a virus like no other or, as Neville Hodgkinson expressed it two decades ago, ”a virus that never was”.298
The first study to define the relationship between AIDS and sexual activity in homosexual men was published in two papers in 1982 and 1984 by Michael Marmor, Alvin Friedman-Kien and their colleagues. In their second, updated study they concluded “Stepwise logistic regression analysis indicated that the number of partners per month in receptive anal-genital intercourse with ejaculation, the number of occasions of "fisting", and cytomegalovirus antibody titers were the only independent and statistically significant variables for discriminating patients from controls".299, 300
That only the passive partner is at risk of AIDS is confirmed in the two longest, largest and best designed studies – the MACS and the Amsterdam Cohort Study which also began in 1984.301. The MACS also documented that "greater sexual activity [receptive anal intercourse] following establishment of HIV-1 infection leads to exposure to promoters or co-factors that augment (or determine) the rate of progression to AIDS"302 (emphasis added). In other words, by factors other than HIV. If such factors act after HIV infection then the same “non-HIV” factors must also act before infection. Indeed, the Amsterdam study showed that both immune suppression (decrease in T4 cells) and immune activation (which is currently accepted to be the cause of the clinical syndrome) precede HIV infection.301 The fact that the only sexual act directly related to the development of AIDS is “receptive anal-genital intercourse with ejaculation” means that the cause must be a non-infectious agent(s) present in semen or semen itself.
At the beginning of the AIDS era epidemiologists had an understandable leaning towards an infectious origin for AIDS. This has continued to the present, which means our hypothesis that semen itself plays a role in AIDS remains largely unexamined. Epidemiological studies to test this hypothesis require cohorts of men and women devoid of other AIDS risk factors who have high frequencies of ejaculatory passive anal intercourse with HIV-negative men. Unfortunately, because of the substantive bias towards HIV, no studies of this nature have been published. However, there is at least one other type of study that could provide supportive evidence.
If the cumulative quantity of anally received semen is the cause of a positive antibody test and AIDS, rather than an infectious agent present in semen, then the number of episodes of passive anal sex with ejaculation will prove a greater risk than the number of sexual partners. Epidemiologists have had many opportunities to report these data but we could find only one such study involving homosexual men.
44
In 1985 Janet Nicholson, Harold Jaffe and their colleagues at the CDC reported that "In the year before testing, homosexual men who were seropositive tended to have a greater number of sexual partners (p = 0.009), more episodes of receptive anal intercourse (p < 0.001), and more frequent active (p < 0.001) and receptive (p = 0.023) insertion of hands into the rectum...The number of episodes of receptive anal intercourse per year was the variable most highly associated with HTLV-III/LAV [HIV] seropositivity (F = 27. p < 0.001). After adjustment for this variable, no other variable was statistically significant", and in a subgroup of men analysed, the quantity of semen was the only significant risk factor.303
Nancy Padian’s group reported the same findings in a cross-sectional study of “Male-to-Female Transmission of Human Immunodeficiency Virus”:296 "Ninety-seven female sexual partners of 93 men infected with human immunodeficiency virus were studied...23% of the women were infected...Anal intercourse significantly discriminated between seronegative and seropositive women...The number of sexual contacts (whether vaginal, anal or oral), was significantly associated with infection...whereas general sexual activity (as measured by number of sexual partners [median 2.5 for seropositive; 4 for seronegative women] and number of sexually transmitted diseases) was not associated with HIV infection". The facts that anal intercourse and the number of sexual contacts were the discriminating factors for seropositivity show that homosexual men and heterosexual women share the same sexual risk factors for AIDS.
Epidemiologically the relationship between sex and AIDS is the same as that between sex and pregnancy. In other words, like pregnancy, AIDS and a positive antibody test can be sexually acquired but not sexually transmitted. However, while pregnancy can be acquired by a single act of sexual intercourse, for AIDS to appear a high frequency of receptive anal intercourse over a long period is necessary. This is facilitated by the facts that (a) in contrast to intra-vaginal ejaculation, anally deposited semen is retained and absorbed; (b) the vasculature of the rectum is separated from the lumen by a single layer of absorptive cells compared to the multilayered, skin-like lining of the vagina.
That exposure to semen is causally related to AIDS is supported by additional evidence including the following:
- Semen is the best known biological mitogen and one of the most potent biological oxidants.273, 304
- HIV-positive and AIDS patients are oxidised.278, 279
-
Theoretical and experimental evidence has long existed that semen is carcinogenic and
immunosuppressive.273, 277, 305-307 -
Rectal and colonic trauma accompanying passive anal intercourse facilitate the absorption
of semen and are proven risk factors. -
The use of volatile nitrites is a risk factor and such agents cause smooth muscle relaxation
and vasodilation which may also facilitate absorption of semen. - Nitrites are also oxidising agents and may act synergistically with semen.
The evidence shows that AIDS is not a disease of sexual orientation but of sexual practices, passive anal intercourse in men and women; and "anal intercourse may be practiced by a much larger absolute population of heterosexuals than of homosexuals".308 It is not the sexual act per se but high frequencies of passive anal intercourse with ejaculation combined with drug use and trauma to the intestinal lining which facilitate systemic absorption of semen and other toxins.309-311 If homosexual men had not been exposed to relatively larger quantities of semen compared to heterosexual women in the late 1970s, the systemic effects of semen, like their local effects in cervical and anal cancer,277, 305, 306, 312 might never have come to notice as a new phenomenon. By accepting and promoting the HIV theory the homosexual community may ultimately do more harm than good to their cause and themselves.313, 314 See also our 2010 manuscript, AIDS - Sexually transmitted or sexually acquired?315 HERE
45
WHERE WE ARE NOW
Numerous commentators affirm that since the introduction of highly active antiretroviral drug combination therapies (HAART) in 1996, AIDS has been transformed from an early and certain death into a “manageable chronic condition”. For example, the UK based AIDS charity AVERT states “The history of the HIV and AIDS epidemic began in illness, fear and death. However, the development of highly effective antiretroviral drugs represented a major turning point by allowing people living with HIV to live long and healthy lives”.316 Jacqueline Jourjy from the University of Florida writes, “Highly active antiretroviral therapy (ART) and its widespread availability have revolutionized the landscape of HIV care and patient outcomes, transforming infection with HIV into a manageable chronic condition rather than a life-limiting disease”.317 Joseph Sonnabend praises antiretroviral drugs as a “wonderful blessing”,318 evoking the 19th century American surgeon John Warren’s prophecy concerning anaesthesia – “a blessing to mankind”.
In the view of the recently retired “AIDSTruth team”, “The efficacy of antiretrovirals, starting with AZT in 1987, is unequivocal...By 1996 the benefits of triple-drug antiretroviral treatment were profound. And in the past month [July/August 2015], the publication of the START and TEMPRANO studies has shown yet again, in randomised clinical trial conditions, how effective these medicines are at keeping people with HIV healthy”.319 In their 2016 online valedictory, the year that marks a decade since the introduction of HAART, the AIDSTruth team declared, “Our work is done...HIV was proven to be the cause of AIDS in 1984. By 1987 there was no reasonable doubt...AIDSTruth began in 2006 to provide accurate information that countered the nonsense of AIDS denialism. We have long since reached the point where we—the people who have in one way or another been involved in running this website—believe that AIDS denialism died as an effective political force. We have therefore decided that there are no further compelling reasons
to continue updating this website”.320
Even though the HIV theory was considered proven by 1984, the success attributed to the modern antiretroviral drug regimes is endorsed as another “nail in the coffin” of those who question the HIV theory. In 2015 the immunologist and Nobel laureate Peter Doherty wrote: “Fortunately, that situation changed when the chemists provided the next nail in the coffin of the idea that HIV does not cause AIDS, the development of ‘designer’ drugs that specifically target the virus and block HIV replication...In fact most of the serious HIV scientist sceptics backed off when it became obvious that the anti-retroviral drugs work so effectively to allow those who are infected to live reasonably normal lives”.321
HIV experts assert that the pharmacological basis of HAART is the ability of combinations of “antiretroviral agents to control HIV replication”322 which results in their clinical benefits.222 Let us assume the benefits of HAART are profound323 and their harm is zero.324 What is the evidence that the effect of HAART on morbidity and mortality accrue from an anti-HIV-replication effect and such benefits prove HIV exists and is the cause of AIDS?
The drug with only one pharmacological effect awaits invention. Their multiple effects are affirmed not only by drug toxicities (“side-effects”) but by numerous other actions. A google search reveals the multiple properties and uses of aspirin and statins for example.325 There is also the recent phenomenon of drug “Repositioning, reprofiling, repurposing. Whatever you call it, finding new careers for old drugs is fast becoming big business. Considerable revenues and savings are there to be made in discovering alternative purposes for known compounds”.326, 327 For example, there are over 80 drugs, including antihistamines, calcium channel blockers, antidepressants and benztropine, reported to protect cells against Ebola virus infection.328, 329 The Repurposing Drugs in Oncology (ReDO) Project, an international consortium which oversees such drug discovery for cancer patients,330 recently published a review “that [the antihypertensive, anti-anxiety and cardiac drug] propranolol has potent anti-cancer effects, as evidenced by in vitro, in vivo and a range of clinical data”.331
46
Antiretroviral drugs are no exception to the rule. Regardless of putative “anti-HIV” effects, antiretroviral drugs are toxic to microbes that cause some of the most common and severe AIDS defining diseases, such as tuberculosis and yeast infections. In a 2007 review of “Inhibitors of HIV- 1 Protease” the authors concluded, “many recent reports showed this class of drugs to be effective as antitumor agents, as apoptosis enhancers, antibacterials (for example against Mycobacterium tuberculosis infection), antifungals (for example against Candida albicans), antimalarials, antiSARS and anti-influenza agents. The biochemical/physiological mechanisms underlying such non- antiviral effects also started to be understood ultimately. Such findings open novel possibilities for the design of pharmacological agents useful not only for managing viral infections, but many other diffuse diseases, such as tumors, bacterial, fungal and protozoa (malaria) infections”.332 In a 2009 Lancet Oncology review oncologist Warren Chow wrote, “The use of anti-HIV drugs as cancer treatments is not new...HIV protease inhibitors are thus a new class of anticancer drugs with multiple effects, and other anti-HIV drugs might hold similar promise”.333 In May 2016 Kyle Anderson and colleagues reported that the reverse transcriptase inhibitor “anti-HIV medication” Efavirenz activates an enzyme that controls cholesterol elimination from the brain and concluded, it “has a strong potential as a new anti-Alzheimer disease therapy”.334 The aim of ARVs is to prevent the onset of AIDS indicator diseases in HIV-positive individuals. Hence, there is ample scope for them to be a “wonderful blessing” regardless of what effect they may or may not have on “HIV” replication. And unless the currently antiretroviral drug combinations are drugs like no others, and indeed have only one effect, that is, they inhibit HIV replication to the exclusion of all else, it is scientifically impossible to claim their clinical benefits prove AIDS is caused by a retrovirus HIV.335
The TEMPRANO study336 is cited by the AIDSTruth team in support of the HIV theory. In this paper the authors treated HIV-positive individuals, including patients with normal T4 cells counts (≥ 500 cells/ml), with ART alone or ART plus isoniazid (IPT). (Isoniazid is antibacterial and used in the prevention and treatment of tuberculosis). They found that “ART and 6 months of IPT independently led to lower rates...of severe HIV-related illness”. If a non-antiretroviral compound is capable of benefiting severe HIV-related illness, on what basis can one assert “HAART” drug combinations are specifically antiretroviral?
CDC Graph 1
47
CDC Graph 2
The first graph published by the Centers for Disease Control plots the estimated incidence and deaths from AIDS per quarter from the beginning of 1985 until June 1997.337 The second graph plots AIDS deaths 1987-2010 in the more familiar form of annual data (intercepts added).
When interpreting these data several historical caveats should be borne in mind:
- In June 1981, soon after receiving information of a sudden and rapid increase in the incidence of KS and PCP in homosexual men, the CDC assembled a task force of 32 mainly physicians to actively survey 18 major US institutions for these diseases.338
- The CDC reported 159 cases of AIDS between June and November 1981 of which 149 (94%) had KS or PCP or both. The overall mortality rate was 38% and 61% for PCP.
- In late September 1981 the CDC reported that in the ensuing 11 months there were 593 cases with 243 deaths (41%) and noted that “the [mortality] rate exceeds 60% for cases diagnosed over a year ago”.
- The CDC published the first AIDS surveillance definition in 1982. AIDS was defined as “illnesses in a person who 1) has either biopsy proven KS or biopsy – or culture-proven life threatening opportunistic infection, 2) is under 60, and 3) has no history of either immunosuppressive underlying illness or immunosuppressive therapy”.339, 340
- For all practical purposes, prior to 1985 AIDS consisted of KS or PCP or both.
- The introduction of “mild and moderate disease” as indicating AIDS, which commenced in the last quarter of 1984, coincided with the acceptance of HIV as the cause of AIDS in all risk groups and the first redefinition of AIDS by the CDC.69, 341 As in 1982, the 1985 definition mandated a definitive diagnosis of AIDS indicator diseases using investigative procedures that included biopsy, cytology and microbial cultures.
- The 1985 definition also introduced HIV antibody testing. However, “In the absence of test results, patients satisfying all other criteria in this definition are included as [AIDS] cases”.342
-
The New York State Health Department found that by the beginning of 1987, 13% of 1329 reported AIDS cases with a positive HIV antibody test, clinically had symptoms suggestive
48
of AIDS diseases but these were not definitively diagnosed. A similar study found that approximately 11% of cases had a presumptive diagnosis because, according to one AIDS epidemiologist “Many physicians are familiar enough with AIDS now that when they see a young man with pneumonia, they can make a reasonable presumptive diagnosis [of PCP] without resorting to [open lung] biopsy”343
- Because of non-compliance, that is, physicians reporting cases not meeting the 1982 or 1985 case definitions, the CDC accepted that the 1985 definition made it “unnecessarily difficult to diagnose” AIDS and thus underestimated the true number of cases. This led to the third, 1987 CDC AIDS definition, which increased the number of indicator diseases from 6 to 23, as well as legitimatising the reporting of AIDS without a definitive diagnosis of some AIDS indicator diseases (“Presumptive diagnoses are accepted”), and without evidence of immune deficiency. More importantly, although the definition considered HIV to be the sole cause of AIDS, individuals were to be reported when evidence of HIV infection was “not performed or gave inconclusive results”, or even “With laboratory test results negative for HIV infection”.344
- In 1993 the AIDS definition was revised a third time adding another three indicator diseases (pulmonary tuberculosis, recurrent pneumonia and invasive cervical cancer345). Most significantly, under this definition a person with a T4 cell count < 200 cells/mm3 (normal range 500-1200 cells/mm3) had AIDS even in the absence of an indicator disease. This obliged physicians to report as AIDS, for example, both a clinically healthy HIV-positive individual with a low T4 cell count and a patient dying from PCP (who might have a count > 500 cells/mm3).
- The “1993 definition implementation” induced a substantial spike in the annual AIDS incidence compared to the 1987 incidence. For example, in a study of 532 HIV-positive individuals attending the City Hospital, Edinburgh, up to the end of July 1991, researchers determined that the changed definition caused a doubling of AIDS cases if based on the first of two consecutive CD4 cell counts ≤ 200 cells/mm3 and a trebling if based on one count ≤ 200 cells/mm3.346
- The first HIV antibody screening [EIA, ELISA] assays were licensed by the Food and Drug Administration (FDA) in March 1985. Repeatedly reactive screening assays were to be “confirmed” by the Western blot. The criteria for a positive Western blot were the presence of a p24 or gp41 band, or both. Between 1985 and late 1987 Western blots were performed by one of several means: “"home brew" methods, outside reference laboratory testing commonly offered by the ELISA assay manufacturer, or more commonly, with non- licensed western blot kits”.316, 347
- Between 1985-1987 it became apparent that 15-40% individuals not at risk of AIDS could have one or two Western blots bands, commonly p24.127, 348-351
- In late 1987 the FDA licensed the first HIV Western blot kit, adding bands and mandating that the interpretive criteria include at least two bands. Hence, the process of selecting particular band patterns minimised the probability that low risk individuals, (those not in an AIDS risk group – homosexuals, drugs users, haemophiliacs, Africans), would be classified HIV-positive. Significantly, no band patterns have been validated against HIV itself, that is, HIV isolation/purification.110 Nonetheless, even now the interpretive criteria are not standardised because, since 1987, the number and positions of bands defining a positive test have progressively changed and vary between countries, institutions and laboratories. This means that one and the same person may be HIV-positive under one jurisdiction but not under another.110, 111, 117, 352
- Since HIV antibody testing was not available between 1981 and 1985, at the time of diagnosis and reporting, the HIV status of all AIDS cases was unknown.
- Likewise, the infection status of an unknown number of living or deceased patients diagnosed according to the pre-1987 Western blot criteria is unknown.
49
These should
data, including the increasing number of deaths from AIDS, peaking at the end of 1994, be interpreted in light of the above and the following:
- The text accompanying the CDC December 1997 HIV/AIDS surveillance (first) graph states “The top curve represents the number of cases diagnosed with AIDS using the 1993 definition criteria after adjustments for reporting delays. It represents the distribution of all cases diagnosed with AIDS and illustrates the distorting effect of the change in the [1993] case definition” (emphasis added). In fact the distortion is not solely the result of the 1993 definition but also the 1985 and most significantly the 1987 definitions. And, as the graph confirms, the “Estimated incidence of AIDS” began to decrease when the AIDS surveillance redefinitions ceased.
- The text also affirms “the bottom curve which represents the reported number of deaths of persons with AIDS [uses] the 1993 case definition after adjustments for reporting delays”. However, “Reported deaths are not necessarily caused by HIV-related disease”. In other words, “AIDS deaths” are not necessarily “deaths from AIDS”.
- According to Harry Haverkos from The National Institute on Drug Abuse, “the percentage of homosexual AIDS patients with KS has declined during the past 6 years [1984-1990]... Possible reasons for the decline include changes in homosexual behaviors, leading to the practice of safer sexual techniques, and a decrease in the use of nitrite inhalants”.353, 354 In 1996 Gregory Dore and his colleagues published Australian data showing that “The percentage of people who developed KS by year of AIDS diagnosis declined from 42.9% in 1983-1984 to 20.8% in 1994 (P< 0.0001). This decline is due to a decreased percentage of people developing KS as their initial AIDS illness (35.7% in 1983-1984 to 13.4% in 1994; P < 0.0001).355 In 1990 Valerie Beral and her colleagues noted similar reductions.356 These data, combined with the high case-fatality rate for KS during the early AIDS era, suggest that AIDS, at least in the major risk group, was beginning to wane even before the first, 1985, AIDS redefinition.
- By 1990 HIV experts accepted that KS is not caused directly (by HIV infection) or indirectly (by immune deficiency).356 Yet KS, the principal reason for the retroviral AIDS hypothesis, remains an AIDS indicator disease. This not only makes no sense, it also distorts the interpretation of case data.277
Hence, for many reasons including:
1.
2.
3.
Repeated redefinitions of AIDS with the addition of an increasing number of indicator diseases which could be:
(a) “mild and moderate”; (b) non-definitively diagnosed; (c) not caused by HIV or immune deficiency (KS).
Defining a case of AIDS: (a) in the absence of an indicator disease; (b) without tests for immune deficiency; (c) in the absence of HIV antibody testing; (d) with negative tests for HIV.
The inclusion of clinically healthy individuals,
an increasing number of AIDS cases and a decreasing number of deaths and death rate from AIDS is to be expected. The third CDC graph confirms that this is the case.
50
CDC Graph 3
In the text accompanying this graph the CDC writes, “The decrease in the rate of death among persons with infection classified as stage 3 [AIDS] during 1987—1995 was due partly to antiretroviral therapy less potent than highly active antiretroviral therapy (HAART), which was introduced in 1996 (e.g., monotherapy with zidovudine, or combination therapy with two nucleoside reverse transcriptase inhibitors [NRTIs] but no protease inhibitor). The rapid decrease in the death rate in 1996 and 1997 was largely due to the use of HAART”.
However:
- According to John Bartlett, Director, HIV Care Program, Johns Hopkins Hospital, Baltimore, “The 13th Conference on Retroviruses and Opportunistic Infections, held in Denver, Colorado, February 5-8, 2006, marks the beginning of the year that is regarded as the 10-year anniversary of the introduction of highly active (triple-drug) antiretroviral therapy (HAART)”.357 “Introduction” does not equate to widespread or universal use, even in one country. In 2015 Mira Hleyhel and her colleagues in France and the United Kingdom published a review of treatments of 6138 HIV-1-infected individuals since "the advent of combination antiretroviral therapy (cART) in 1996". They classified cART into “pre-cART (1992-1996) and early cART (1997-2000)”.358
- In the second CDC graph, the “rapid decline” in the number of AIDS deaths began at the end of 1994, two years before the introduction of HAART. During the pre-HAART year of 1995 the decline in AIDS death rate accelerated while the following year, when HAART was introduced, the decline in the death rate continued at the same rate. In other words, “The rapid decrease in the death rate in 1996 and 1997 [that] was largely due to the use of HAART” was in place before the introduction of HAART and HAART did not affect the rate of decline.
-
The third graph shows that the annual death rate from AIDS amongst AIDS patients has been declining since 1987. This cannot be due to zidovudine monotherapy because (a) AZT was not FDA approved for general use until the end of 1987; (b) like HAART its use was not universal at the time of approval; (c) its use was limited by its high dose, difficult dosing schedule and toxicities; and, (d) AZT is administered as an inactive pro-drug which must be tri-phosphorylated intracellularly into its active form. However, the intracellular
51
concentration of tri-phosphorylated AZT is at least one order of magnitude below that required to exert its putative antiretroviral effect.359
- The magnitude of the declining death rate has not been maintained after 1997.
- Attributing the decline in death rate “partly” and “largely” due to ARVs acknowledges the existence of factors unrelated to ARVs and HAART in this decline. This has been argued by others (see below) and that they are substantial is supported by the falling number of deaths and death rate that began before the introduction of HAART.
In addressing the decreasing “Mortality and changing patterns of causes of death” in AIDS patients it is difficult to find a study in which the data have been better collected and analysed than that of Rainer Weber and his colleagues in Switzerland. In 2013 these authors published a paper in HIV Medicine, the journal of the British HIV Association.360
As reported in their figures (a) and (b), amongst the 16,134 participants in the Swiss HIV cohort study the annual “AIDS-associated mortality peaked in 1993” followed by a sharp drop which had reached a 40% reduction at the time HAART was introduced at the beginning of 1996.
A falling death rate before the introduction of HAART can be explained on the basis of:
- As soon as physicians began diagnosing high frequencies of KS and PCP in the late 1970s homosexual men were the first to recognise the relationship between the two diseases and sexual activity/drug use. Many if not the majority, took the necessary precautions298 (“behaviour change”) to decrease their "exposure to promoters or co-factors that augment (or determine) the rate of progression to AIDS".302 Those heavily and repeatedly exposed to these factors in the 1970s would have died during the 1980s.
- As one infectious physician pointed out, "the improved skills of physicians in treating opportunistic infections (and better protocols)", would have resulted in far fewer “AIDS- associated” deaths by the early 1990s (Fabio Franchi, personal communication).
-
This view is shared by Mary Ann Chiasson, assistant commissioner of the New York City Department of Health, speaking at the 4th Conference on Retroviruses and Opportunistic Infections, 22-26 January, 1997. Reporting Chiasson in Science Magazine’s Research News a week after the conference, staff writer Jon Cohen wrote, “In New York City, which accounts for 16% of U.S. AIDS cases, AIDS deaths last year dropped by 30%. But health officials did not attribute the drop to increased use of protease Inhibitors. According to Chiasson, the AIDS death rate began to fall before the two most powerful drugs reached the market last spring [March-June 1996]. She suggested that the decline in deaths may instead be linked more closely to an increase in federal funding in 1994 for AIDS patients, which led to better prevention and treatment of opportunistic infections”.361
52
4. A declining death rate is the expected outcome of introducing “mild and moderate” diseases into the AIDS surveillance definitions.
In view of all these factors it is highly unlikely we will ever know how the incidence of “KS/PCP AIDS” may have evolved following its inception as a new syndrome that devastated homosexual men during the 1980s. AIDS as defined in the 1982 definition may have declined to a level determined by the heightened awareness that arose as a result of the high frequencies of KS and PCP reported at the beginning of the AIDS era.355, 356, 362-365 The reason for the contrary appears unrelated to the natural history of a retroviral infection. Rather, it is a result of many artefactual factors especially definitional changes, most importantly those that occurred in 1987 and 1993.
The Weber et al graphical data also show that between 2002 and 2010 the most frequent causes of death were non-AIDS-related malignancies and liver failure. These conditions are induced by oxidation.208, 223, 272, 275, 366 As Montagnier acknowledges, HAART causes oxidation.65, 219
Nonetheless, the HIV experts, such as members of the AIDSTruth Team, claim it was the introduction of antiretroviral drugs including Azidothymidine (AZT335) in 1987 that led to a decline in the prevalence of AIDS. This claim was made despite the facts that:
- AZT is administered as an inactive pro-drug which must be tri-phosphorylated intracellularly into its active form. However, the concentration of tri-phosphorylated AZT is at least one order of magnitude below that required to exert its putative antiretroviral effect.359
- AZT therapy was introduced in 1987 and by 1990 homosexual men realised that "AZT's alleged benefits are not backed up by hard data, and are not sufficient to compensate for the drug's known toxicities. Recovery from AIDS will come by strengthening the body, not by poisoning it".367-369 In fact it was its toxicities that led John Lauritsen to invent the epithet "Poison by Prescription". Clinical data show that AZT is detrimental to patients.359, 367, 370-373 This is recognised by the HIV experts and their supporters, such as the AIDSTruth Team members, who described AZT toxicities including deaths, as “bumps in the early years of treatment”.319, 374
HAART – the “unexpected” anti-HIV outcome
A theory is as good as its predictions and there are no more pressing predictions for a disease theory than its clinical predictions. Indeed, the rush at the beginning of the 1980s to propose a theory of AIDS reflected the urgency for treatment and prevention.
Two theories were put forward – our cellular redox theory and the HIV theory. The principal clinical predictions of the redox theory were: (a) AIDS patients and those at risk will be oxidised – in particular the sulphydryl groups (SH groups) of the cellular, acid-soluble proteins; and,(b) AIDS can be prevented and treated by the use of antioxidants, especially SH containing compounds, and mild hyperthermia.273, 274, 276, 304, 375 One year after the publication of this theory researchers from Germany proved our first prediction, that is, the SH of the acid-soluble proteins in AIDS patients and those at risk are indeed oxidised. The first and to date the only study that proves our second prediction was conducted by a group of researchers at Stanford University.
The main clinical prediction of the HIV theory of AIDS is: AIDS can be prevented and treated by antiretroviral drugs, HAART. If HIV is the cause of AIDS and the drugs prevent viral replication this will obviously be the case. It is accepted that HAART does not destroy virus particles either free or in infected cells. Rather HAART is said to interrupt the viral replication cycle thereby preventing the production of new virus particles and the subsequent infection of naïve cells. HAART consists of various combinations of reverse transcriptase inhibitor (RTI) and protease inhibitor (PI) drugs. RTIs prevent infection by inhibiting reverse transcription of the viral RNA into proviral DNA. PIs act on infected cells to prevent the cleavage of “HIV” polyproteins into the smaller proteins required for the assembly of mature, replication competent particles. Given data published in Nature in 1995,
53
that the “lifespan of plasma virus and virus-producing cells is remarkably short (half-life ~2 days”88), the synergistic action of PIs and RTIs should result in a decline in cellular proviral DNA (“viral burden”) in a matter of days as infected cells disappear. Following this there should be a decline in the number of viral particles, “viraemia” (“viral load”) in the plasma. The decline in HIV is said to reverse the decline in cellular immunity (the falling T4 cell count) and thus progression to the clinical syndrome, AIDS/death.
However, according to evidence published by Zaunders and colleagues, the decline in “viral load” is not preceded or accompanied by a decrease in proviral DNA. To the contrary. Their data show that HAART decreases the “viral load” “from 6.0 log10 copies/mL at baseline to <400 copies/mL with 20 weeks and was <50 copies/mL in 13/13 patients after week 36” while at the same time HAART “had little direct effect on HIV-1 DNA burden [proviral DNA]”.376, 377 In other words, the “viral load”, the “HIV” RNA, declines despite the fact that the viral progenitor, the proviral DNA, remains unchanged. This means that by whatever mechanism HAART reduces the “viral load”, it cannot be by inhibition of reverse transcription of RNA or the assembly of viral particles. Either “HIV” contravenes the theory of the retroviral replication cycle, or “HIV” RNA or DNA or both is/are not retroviral; and there is no relationship between them. This is supported by evidence presented 8 years earlier at the VIIth International AIDS Conference by Mikovits and colleagues. These authors showed that monocytes from HIV-positive patients in which no HIV DNA could be detected, even by the polymerase chain reaction, became positive for HIV RNA after co-cultivation with normal ConA activated T cells (ConA is a mitogen, oxidising agent). Mikovits et al concluded that "HIV expression [its RNA genome] can be activated in monocytes which lack detectable HIV DNA".378. This supports our claim that the “HIV” RNA is not retroviral-related and is either an edited cellular RNA or novel RNA.
The AIDSTruth team also cited the START study (Strategic Timing of Antiretroviral Therapy) as further proof that HIV is the cause of AIDS.379 The START authors began their paper asserting the HIV theory of AIDS: “The immune compromise caused by the human immunodeficiency virus (HIV) is characterised by a loss of CD4+ T cells. Rates of HIV-associated complications and death increase as the number of these cells in peripheral blood (CD4+ count) declines”. Indeed, given the claim that initial HIV infection of an individual instigates abundant HIV replication from inception until death, and that the life of infected T-cells is only a few days, the administration of ARV drug combinations should lead both in vitro and in vivo to (i) decreased formation of proviral DNA; and thus (ii) decreased viral load; and ultimately (iii) increased CD4 cells; and, (iv) a decrease in AIDS/death; and with all these parameters highly correlated.
However, the evidence they published contradicts their claim. In regard to the relationship between HIV viremia, CD4 cell count and their primary end points of death/AIDS the authors state: “The risk of AIDS was not zero among patients receiving antiretroviral therapy, even among those who had full viral suppression while receiving antiretroviral drugs”. Furthermore, “Most of the AIDS-related and non-AIDS related events occurred when patients had a high CD4+ count...with the latest CD4+ count of more than 500 cells per cubic millimetre, which may indicate that a substantial part of the beneficial effect of immediate treatment is due to changes induced by antiretroviral therapy in markers other than the CD4+ count. This finding is consistent with the results of studies that were used as a basis for our study and with other large observational studies conducted both in resource-rich and resource-poor settings...It further supports the need for better markers of impaired immune function and research on treatments to use along with antiretroviral therapy to reduce disease among HIV-positive patients” (emphasis added). In other words, not only do their data fail to support the HIV theory, the authors themselves cite clinical trials conducted during the HAART era that arrive at the same conclusion.
The START findings were also reached in an earlier, 2010 study on antiretroviral therapy conducted in Uganda.380 Researchers from Uganda, UK and Zimbabwe recruited 600 patients of whom 563 completed 48 weeks treatment. In an analysis of the relationship between HIV (“viral load”), CD4 count, and clinical outcome, they found “a disconnect between
54
immunological/virological responses and clinical outcome at 48 weeks”, for which they could find “no explanation”. The authors concluded: “Given the universal use of CD4 and viral load for the assessment of ART effectiveness in clinical trials, our unexpected findings are of concern”. The authors have reason to be concerned. Their findings mean either the trial was badly designed and/or executed or the HIV theory of AIDS is wrong. The latter should have been apparent because the authors themselves cite several studies that confirm their findings. This included one study that showed an inverse relationship between virological/immunological responses and clinical outcome.
Among those cited is a paper by the epidemiologist Edward Mills and five colleagues in public health and infectious diseases from Canada, the United Kingdom and the USA. In 2008 they published “A Review In Antiretroviral Research” in HIV Medicine, the journal of the British HIV Association.381 The review is “a meta-regression analysis” of data obtained from 178 (the best out of 10,372 initially identified), randomised clinical trials (RCTs) to determine:
- The effect HAART has on two “surrogate markers”382 – “HIV-1 RNA viral load” and CD4 counts.
- “The effectiveness of CD4 T-cell changes and viral load thresholds in predicting progression to AIDS/death”.
The authors state, “Our study represents the largest assessment of the relationship between surrogate outcomes and clinical events to date” and the 178 RCTs that lasted between “48-96 weeks”. They reported that “most HAART therapies appear to offer high levels of CD4 and viral load control” but they “were unable to demonstrate a relationship between change in CD4 cell count or viral load and clinical events”. Even if one assumes there are beneficial effects from HAART therapy, the fact that no correlation exists between virological and clinical outcomes means the benefits are not the result of an antiretroviral effect.383
Most importantly, HIV viremia and CD4 (T4) cell counts are not just any surrogate markers. According to the HIV theory they are the cause and underlying mechanism for the clinical syndrome, AIDS/death. These surrogate markers are the HIV theory of AIDS: HIV low T4s (immune deficiency) AIDS/death. If the best randomised controlled trials to date, lasting between one and two years, show no “relationship between change in CD4 cell count or viral load and clinical events” then either (a) the RNA is not HIV RNA, that is, there is no HIV genome, and if there is no HIV genome there is no virus to cause AIDS; or, (b) the RNA is HIV RNA but HIV is not the cause of AIDS. Since to date nobody has published proof that what was assumed to be the “HIV RNA” originates from a retroviral particle, the former explanation - that there is no virus - must hold true.
Conclusion
On the basis of the presently available data in the scientific literature, one has no choice but to conclude that whatever “HIV” is, it is not “the virus that causes AIDS”, or even “a real virus”.
Eleni Papadopulos-Eleopulos, Valendar F. Turner, John M Papadimitriou, Barry A. P. Page, David Causer
Posted online July 12th 2017. LINK
55
Explanatory note
On 21st February 2017 the Perth Group emailed this manuscript to Nature, The Lancet, the British Medical Journal, Science, Medical Hypotheses, the Journal of the American Medical Association and the New England Journal of Medicine.
The following covering note was addressed to each editor-in-chief.
My colleagues and I have a somewhat unusual request of your editorial office. The attached document is a detailed re-evaluation of the HIV/AIDS theory. For several decades we have closely followed the evolution of this theory and the data upon which it was based. In our view the theory was formulated on evidence and observations that were not always subjected to the expected strict scientific rigour. Consequently its conclusions and predictions need to be thoroughly questioned and the precise nature of "HIV" redefined.
We are aware that the topic is contentious, especially as some well-publicised challenges to the orthodox view have had deleterious public health implications.
Our request to you as guardians of scientific thought and integrity is to assess our critique, and see whether in your considered opinion it is worthy of being brought to the attention of the scientific community. If you decide that it is, we would then like to prepare, under your guidance, a concise version for publication.
Despite repeated requests, three editors did not acknowledge receipt of our email. All responses came from editorial committees. One was “we do not encourage pre-submission enquiries”; another, we “respectfully pass on having the opportunity to publish a paper on this topic at this time. Best of luck publishing your work in another good journal”. A third, “After considering its focus, content, and interest, we made the editorial decision not to consider your proposal further. We are informing you of this decision promptly so that you can submit your manuscript elsewhere”. All replies ignored our request for a private appraisal and/or responded as if the manuscript were a submission for publication (a futile exercise given that no editor would accept a paper of this length).
56
References and Notes
1. CDC. Prevention. Pneumocystis pneumonia-Los Angeles. Morbidity and Mortality Weekly Report 1981. 30:250-252. 2. Friedman-Kien A, Laubenstein L, Marmor M, Hymes K, Green J, Ragaz A, Gottlieb J, Muggia F, Demopoulos R, Weintraub M. Kaposi's sarcoma and Pneumocystis pneumonia among homosexual men--New York City and California. Morbidity and Mortality Weekly Report 1981. 30:305- 308. 3. It is a common belief that the homosexual men who ushered in the AIDS era in the early 1980s were previously healthy. For example the seminal paper by Gottlieb et al has the title "Pneumocystis carinii pneumonia and mucosal candidiasis in previously healthy homosexual men". "Previously healthy" was not the experience of patients under the care of Dr. Joseph Sonnabend, an infectious disease physician working in New York City at the time AIDS began. Nor Michael Callen, also a resident of New York City at that time. Callen's interview with the British science writer Neville Hodgkinson challenges the notion of "previously healthy": "From 1973, when he [Callen] came out as a homosexual, to 1975, he only got mononucleosis and non-specific urethritis (NSU). In 1975 he had his first bout of gonorrhoea. 'Not bad', I [Callen] thought. I'd had maybe 200 different partners and I'd only gotten the clap twice. But from there, it all began to snowball. First came hepatitis A in 1976. Then more NSU and gonorrhoea. In 1977, amoebas [intestinal parasites]-and hepatitis B. More NSU and gonorrhoea. 1978: more amoebas. And my first case of shigella [which causes dysentery]. And of course more VD. Then in 1979, hepatitis a third time: this time non-A, non-B. More amoebas, adding giardias this time. And a fissure. And my first case of syphilis. And of course more gonorrhoea [penile, anal and oral]. In 1980: the usual gonorrhoea, shigella twice, and more amoebas. By 1981, I got some combination of venereal diseases EACH AND EVERY TIME I had sex'. Added to that list were herpes simplex types I and II; venereal warts, salmonella; chlamydia; cytomegalovirus (CMV); Epstein-Barr virus (EBV); mononucleosis; and finally cryptosporidiosis ["I had a disease of cattle!"], and the other infections too numerous to count, which at the time he did not connect to his promiscuous sex life" (capitals in original). Callen died of AIDS in December 1993 aged 38. See: Hodgkinson N. AIDS The failure of contemporary science: How a virus that never was deceived the world. London: Fourth Estate; 1996.
4. Prior to 1993 the CDC definition of AIDS included "disease caused by M. tuberculosis, extrapulmonary (involving at least one site outside the lungs, regardless of whether there is concurrent pulmonary involvement"). Then, after 1993, pulmonary tuberculosis in the absence of disease outside the lungs, became an AIDS indicator disease. This redefined millions of patients with tuberculosis as AIDS patients. The absurdity of such definitional changes was highlighted in Brent Leung's 2009 documentary, House of Numbers.
http://www.cdc.gov/mmwr/pdf/other/mmsu3601.pdf http://www.cdc.gov/mmwr/preview/mmwrhtml/00018871.htm 5. Montagnier L. Virus. New York, WW Norton & Company Inc, 2000. 6. The beginning of HIV can be traced back 45 years to the signing on December 23rd 1971 of The National Cancer Act of 1971 by US President Richard Nixon. Commonly referred to as the War On Cancer it devolved into three streams – epidemiological studies, the development of anti-cancer drugs and the search for cancer causing viruses. Viruses had been implicated in human cancers prior to 1971 but the search intensified as funds controlled by the National Institutes for Health began to flow. The 1970s witnessed a proliferation of virologists, virology laboratories and virology publications but by 1980 the result of a decade of concentrated study was a claim by Robert Gallo to have discovered one human retrovirus. This discovery, later renamed human T-cell lymphotropic virus-I (HTLV-I), is said to cause a rare form of leukaemia involving T4 lymphocytes (adult T-cell leukaemia), although in Harold Varmus’ view, “The HTLVs have been proven to be curious agents, difficult to understand as pathogens and difficult to study as infectious viruses”. If HTLV-I does cause leukaemia its discovery was not about to dent cancer statistics because of its rarity, 140 cases in the US between 1993 and 2008 – rarer than US cases of leprosy. (Contrast this with a new case of breast cancer every three minutes). The first decade of virus hunting brought such little return that as the
57
AIDS era dawned, legislators and the general public were becoming increasingly disillusioned with the cost/benefit of The War. Some virologists were facing unemployment. As a book reviewer at The Economist recently wrote, it was “a piecemeal effort that has been captured, because of flaws in the original legislation, by individual researchers pursuing individual agendas rather than being the goal- directed, co-ordinated project, similar to the moon-shot, that many had in mind back in the 1970s”. By 1980 it was apparent that cancer was not about to surrender to anyone including the virology community. In 2013 Clifton Leaf published a scholarly account detailing his view that The National Cancer Act was doomed from the day it was signed.
See: Varmus H. Retroviruses. Science 1988 240:1427-1435. Chihara D, Ito H, Katanoda K, Shibata A, Matsuda T, Tajima K, Sobue T, Matsuo K. Increase in incidence of adult T-cell leukemia/lymphoma in non-endemic areas of Japan and the United States. Cancer Sci 2012 103:1857-1860. Nolen L, Haberling D, Scollard D, Truman R, Rodriguez-Lainz A, Blum L, Blaney D. Incidence of Hansen's Disease - United States, 1994-2011. MMWR Morbidity and mortality weekly report 2014 63:969-972. http://www.ncbi.nlm.nih.gov/pubmed/25356604 Anonymous. Hard pounding, gentlemen. There is no real secret about how to win the war on cancer. http://www.economist.com/news/books-and-arts/21586279-there-no-real-sec…- cancer-hard-pounding-gentlemen Part III The Cancer Culture. Chapter 8 "The Wrong Bill" in Leaf C. The Truth in Small Doses: Why We're Losing the War on Cancer-and how to Win it. Simon and Schuster, 2013.
7. The delay between an HIV antibody positive diagnosis and AIDS varies up to 50 fold. In untreated, HIV positive individuals the annual AIDS risk is 1-2% for haemophiliacs, 5% for drug users and homosexual men and 50% in blood transfusion recipients. Even if there were an AIDS virus, it would not be sufficient to cause AIDS. As infectious disease physician Joseph Sonnabend pointed out, with the possible exception of rabies not everyone infected with a microbe falls ill. Additional host and/or environmental factors (“co-factors”) always play a role. The admission of co-factors complicates proofs of causation including the possibility that "the virus" is a co-factor, that is, not the main factor. 8. Barré-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet C, Axler-Blin C, Vezinet-Brun F, Rouzioux C, Rozenbaum W, Montagnier L. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 1983. 220:868-871. 9. Gallo RC, Salahuddin SZ, Popovic M, Shearer GM, Kaplan M, Haynes BF, Palker TJ, Redfield R, Oleske J, Safai B, White G, Foster P, Markham PD. Frequent Detection and Isolation of Cytopathic Retroviruses (HTLV-III) from Patients with AIDS and at Risk for AIDS. Science 1984. 224:500-503. 10. Popovic M, Sarngadharan MG, Read E, Gallo RC. Detection, Isolation,and Continuous Production of Cytopathic Retroviruses (HTLV-III) from Patients with AIDS and Pre-AIDS. Science 1984. 224:497-500. 11. Sarngadharan M, G., Popovic M, Bruch L, Schupbach J, Gallo R. Antibodies Reactive to Human T-Lymphotrophic Retroviruses (HTLV-III) in the Serum of Patients with AIDS. Science 1984. 224:506-508. 12. Schupbach J, Popovic M, Gilden RV, Gonda MA, Sarngadharan M, Gallo RC. Serological analysis of a Subgroup of Human T-Lymphotrophic Retroviruses (HTLV-III) Associated with AIDS. Science 1984. 224:503-505. 13. Gallo RC, Sarin PS, Kramarsky B, Salahuddin Z, Markham P, Popovic M. First isolation of HTLV-III. Nature 1986. 321:119. 14. Arya SK, Gallo RC, Hahn BH, Shaw GM, Popovic M, Salahuddin S, Wong-Staal F. Homology of genome of AIDS-associated virus with genomes of human T-cell leukemia viruses. Science 1984. 225:927-930. 15. Hahn BH, Shaw GM, Arya SK, Popovic M, Gallo RC, Wong-Staal F. Molecular cloning and characterization of the HTLV-III virus associated with AIDS. Nature 1984. 312:166-169.
58
16. Shaw GM, Hahn BH, Arya S, Groopman JE, Gallo RC, Wong-Staal F. Molecular characterization of human T-cell leukemia (lymphotropic) virus type III in the acquired immune deficiency syndrome. Science 1984. 226:1165-1171. 17. Coffin J, Haase A, Levy J, Montagnier L, Oroszlan S, Teich N, Temin H, Toyoshima K, Varmus H, Vogt P. What to call the AIDS virus? Nature 1986. 321:10.
18. Bent Leung's video interview with Dr. Joseph Sonnabend. New York City 2006.
http://www.youtube.com/watch?v=6Cd2pknV0Hg&sns=em
19. Feldbaum H, Lee K, Patel P. The national security implications of HIV/AIDS. PLoS Med 2006. 3:e171. http://www.ncbi.nlm.nih.gov/pubmed/16752951 20. Lever AM, Berkhout B. 2008 Nobel prize in Medicine for discoverers of HIV. Retrovirology 2008. 5:91. http://www.biomedcentral.com/content/pdf/1742-4690-5-91.pdf
21. Twitter. 20 mai 1983 : découverte du virus responsable du #sida à l'Institut Pasteur.
https://twitter.com/institutpasteur/status/733597192443727873
22. Vahlne A. A historical reflection on the discovery of human retroviruses. Retrovirology 2009. 6:40. http://www.retrovirology.com/content/6/1/40 23. Immediately after the announcement of the 2008 Prize we emailed and sent printed copy by registered mail to Karolinska extensively documenting our reasons why no scientist including Montagnier and Barré-Sinoussi had discovered a retrovirus HIV. We have no knowledge of whether the correspondence was read or received.
24. In 2006/2007 Andre Chad Parenzee made an application for leave to appeal to his earlier conviction for endangering the lives of three women following "unprotected sexual intercourse...when he knew he was infected with the virus HIV". (R v PARENZEE [2007] SASC 143; 248 LSJS 99, https://jade.io/j/?a=outline&id=8353). The application was heard over several weeks and three sessions October 2006-January 2007 at the Supreme Court of South Australia. During the proceedings the South Australian Director of Public Prosecutions claimed there is no need for purification to prove the existence of a virus. In support of this she submitted the first chapter of a text book, Medical Virology. However, as can be seen by the following quotations from the book, which were read out in court by EPE, the authors of Medical Virology do not support this claim.
"CHEMICAL COMPOSITION OF VIRUSES Methods of Purification An essential prerequisite for the chemical analysis of viruses has been the development of adequate methods of purification. Special problems are created by the close association of viruses with the cells they parasitize; it is not an easy matter to free virions of associated cell debris, or even from viral proteins synthesized in excess in the infected cell... Physical Methods of Purification After partial purification and concentration by chemical methods, or even without any preliminary treatment, virus particles can be separated from soluble contaminants by centrifugation....Equilibrium (isopycnic) [density] gradient centrifugation in dense solutes such as caesium chloride or potassium tartrate (or sucrose in the case of enveloped viruses of low density), on the other hand, separates virions from contaminants according to their buoyant density. After prolonged ultracentrifugation at very high gravitational forces the virions will come to rest in a sharp band in that part of the tube where the solution has the same density as the virions, usually within the range 1.15 – 1.4 [g/ml]." See: White DO, Fenner FJ. Medical Virology. San Diego, Academic Press, 1986, pp. 655. Parenzee Hearing transcripts http://www.tig.org.za/Parenzee_prosecution_transcripts.
25. In a nutshell. Page 37 of the Perth Group's commentary on Brent Leung's documentary The Emperor's New Virus? http://www.youtube.com/watch?v=PQFxratWh7E http://theperthgroup.com/OTHER/ENVCommentary.pdf#page=37 26. HIV experts advise regular monitoring of patients' HIV "viral load". HIV expert Sharon Lewin informs ABC radio Australia listeners "When you start someone on ART [antiretroviral therapy], the virus very rapidly disappears from the blood. In fact, within a month, a person can have over a million copies of the virus per millilitre of blood fall to undetectable levels". "Viral load" and the synonymous term "plasma viraemia" ("virus in the blood") is not, as many physicians,
59
patients and many others imagine, the concentration of viral particles in the bloodstream. The viral load is based on PCR that purports to count the number of "HIV RNA" molecules per millilitre of plasma. (According to Anthony Fauci, "The commercially available RNA detection tests have a sensitivity of 40–80 copies of HIV RNA per milliliter of plasma. Research laboratory–based RNA assays can detect as few as one HIV RNA copy [one molecule] per milliliter"). Since retrovirus particles measure 100 nm in diameter, the only way to determine "copies of the virus" is by electron microscopy. However to date no electron microscopist, including Hans Gelderblom from the Robert Koch Institute in Berlin, has reported the detection of "HIV" particles in the plasma of even one patient with any amount of "viral load". Gelderblom regards this the Holy Grail of HIV research.
27. The difficulty virologists have defining virus isolation can be judged from the following: In his documentary House of Numbers Brent Leung asked Nobel laureate David Baltimore to explain the difference between isolation and purification. Baltimore struggled, was unable to provide a coherent answer, then became visibly exasperated and concluded "Why should I do all of this...This is all textbook stuff you are asking me...I don't want to be your textbook. I've got other things to do". https://www.youtube.com/watch?v=1Li9MO3RfCQ Time: 4:48.
28. Neither virologists nor the scientific literature provides a satisfactory definition of virus isolation. HIV expert Jay Levy defines virus isolation as a "sample of a virus from a defined source", White and Fenner as the ability to "identify a totally unforeseen virus, or even discover an entirely new agent". Montagnier and Weiss as "propagating them [viruses] in cells in culture". The 2013 sixth edition of Fields Virology defines isolation as "Viruses can be isolated from an infected host by harvesting excreted or secreted material, blood, or tissue and testing for induction of the original symptoms in the identical host, or induction of some abnormal pathology in a substitute host, or in a cell culture...Once the presence of a virus has been established, it is often desirable to prepare a genetically pure clone". It goes without saying that if virus isolation is to "take a sample of a virus from a defined source", or "propagating them in cells in culture", one first must have proof the virus exists in "a defined source" or "in cells in culture". Neither is virus isolation "induction of some abnormal pathology" or "once the presence of a virus has been established".
See: Levy JA, Fraenkel-Conrat H, Owens RA. (1994). Virology. 3rd ed. London: Prentice-Hall, 1994. White DO, Fenner FJ. (1994). Medical Virology. 4th ed. San Diego: Academic Press. Turner VF, Weiss R. Email debate with Professor Robin Weiss on the existence of HIV, 1999. www.virusmyth.com/aids/perthgroup/papers2.html http://www.theperthgroup.com/EMAILCORR/vftweiss.html Tahi D. (1998). Did Luc Montagnier discover HIV? Text of video interview with Professor Luc Montagnier at the Pasteur Institute July 18th 1997. Continuum 5:30-34. www.virusmyth.com/aids/data/dtinterviewlm.htm
29. White DO, Fenner FJ. Medical Virology. San Diego, Academic Press, 1986, pp. 655. 30. Levy JA, Fraenkel-Conrat H, Owens RA. Virology. London, Prentice-Hall, 1994, pp. 447. 31. Papadopulos-Eleopulos E, Turner V, Weiss R. Email debate with Professor Robin Weiss on the existence of HIV. 1999. http://www.theperthgroup.com/EMAILCORR/vftweiss.html 32. The Emperor's New Virus? 2012. http://www.youtube.com/watch?v=PQFxratWh7E 33. The Durban declaration. Nature 2000. 406:15-16. http://www.nature.com/nature/journal/v406/n6791/full/406015a0.html 34. Anonymous. The Durban Declaration Signatories. 2000. http://www.nature.com/nature/journal/v406/n6791/suppinfo/406015a0.html 35. Wain-Hobson S. Genesis of the Durban Declaration. 2000. 406:15-16. http://theperthgroup.com/SOUTHAFRICA/whdd.doc 36. "Isolation" invokes the image of the shipwrecked man on a desert island. Indeed island reflects the Latin origin of the word from insulatus – "made into an island”. In 1978 misuse of “isolation” in virology was taken up by CR Madeley, a virologist at the Department of Infectious Diseases, Ruchill Hospital, Glasgow, and CJ Kay, Director of the Historical Thesaurus of English, Department of English Language, University of Glasgow. They suggested the term “recognisate” should be used in place of “isolate” in virology. They argued that “An isolate may be defined as a
60
microorganism grown in pure culture...There is now increasing use of methods for recognising the presence of a microorganism without growing it...To refer to positive results in these tests as “isolates” must be incorrect since they have not been grown...– nor can they be said to be free of other organisms”. In other words, what virologists claim as isolation is not isolation but detection.
Detection means finding evidence of a virus in material which may or may not be pure. In practice it is always a mixture of material. Obviously detection can only be as good as the specificity of the method used for detection. Given that Madeley and Kay were speaking at the 1978 IVth International Conference on Virology it is evident their appeal for correct language went unheeded.
37. Coffin JM, Temin HM. Ribonuclease-sensitive deoxyribonucleic acid polymerase activity in uninfected rat cells and rat cells infected with Rous sarcoma virus. J Virol 1971. 8:630-642. 38. Gallo RC, Wong-Staal F, Reitz M, Gallagher RE, Miller N, Gillespie DH. Some evidence for infectious type-C virus in humans. Edited by Balimore D, Huang AS, Fox CF. New York, Academic Press Inc., 1976, pp. 385-405.
39. Varmus HE. Reverse transcription in bacteria. Cell 1989. 56:721-724. 40. Neurath AR, Strick N, Sproul PSO. Search for hepatitis B virus cell receptors reveals binding sites for interleukin 6 on the virus envelope protein. J Exp Med 1992. 175:461-469. 41. Leung, B. House of Numbers. 2006. http://www.youtube.com/watch?v=BsT4GrimfLQ 42. Pachacz M. No need to be phased. 2001. http://www.theperthgroup.com/POPPAPERS/SharesMagazine2001.pdf 43. Despite all the evidence to the contrary (much of it their own from the 1970s) leading HIV experts still claim reverse transcriptase is retroviral specific. Montagnier's co-worker Jean-Claude Chermann told Djamel Tahi: “The second point is related to the detection of RT activity which is a retrovirus specific enzyme” (D. Tahi. personal communication). 44. Gallo D, Kimpton JS, Dailey P. Comparative studies on use of fresh and frozen peripheral blood lymphocyte specimens for isolation of human immunodeficiency virus and effects of cell lysis on isolation efficiency. J Clin Microbiol 1987. 25:1291-1294. 45. Goudsmit J. Viral Sex-The Nature of AIDS. New York, Oxford University Press, 1997. 46. FRONTLINE. Interview with David Ho. The Age of AIDS. 2006. http://www.pbs.org/wgbh/pages/frontline/aids/interviews/ho.html 47. Tahi D. Did Luc Montagnier discover HIV? Text of video interview with Professor Luc Montagnier at the Pasteur Institute July 18th 1997. Continuum 1998. 5:30-34. http://www.altheal.org/continuum/Vol5no2.pdf http://leederville.net/links/TahiContinuum1998.pdf 48. https://en.wikipedia.org/wiki/Endogenous_retrovirus 49. Lower R, Lower J, Kurth R. The viruses in all of us: Characteristics and biological significance of human endogenous retrovirus sequences. Proceedings of the National Academy of Sciences of the United States of America 1996. 93:5177-5184. 50. Griffiths DJ. Endogenous retroviruses in the human genome sequence. Genome Biol 2001. 2: 1017.1-1017.5. 51. Frank H. Retroviridae. Edited by Nermut MV, Steven AC. Oxford, Elsevier, 1987, pp. 253-256. 52. Todaro GJ, Benveniste RE, Sherr CJ. Interspecies Transfer of RNA Tumour Virus Genes: Implications for the search for "Human" Type C Viruses. Edited by Baltimore D, Huang AS, Fox CS. New York, Academic Press Inc., 1976, pp. 369-384. 53. Weiss RA. Why cell biologists should be aware of genetically transmitted viruses. National Cancer Institute monograph 1978. 183-189. 54. Ono K, Ohashi A, Yamamoto A, Matsukage A, Nishioka N, Takahashi T, Nakayama C, Saneyoshi M. Discrimination of reverse transcriptase from cellular DNA polymerase by kinetic analysis. Cell Mol Biol 1979. 25:323-328. 55. Weissbach A, Baltimore D, Bollum F. Nomenclature of eukaryotic DNA polymerases. Science 1975. 190:401-402. 56. Kaguni LS. DNA polymerase gamma, the mitochondrial replicase. Annu Rev Biochem 2004. 73:293-320. 57. Barré-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet C, Axler-Blin C, Vezinet-Brun F, Rouzioux C, Rozenbaum W, Montagnier L. Electron micrograph 1983. http://leederville.net/links/LMLAV1983.jpg
61
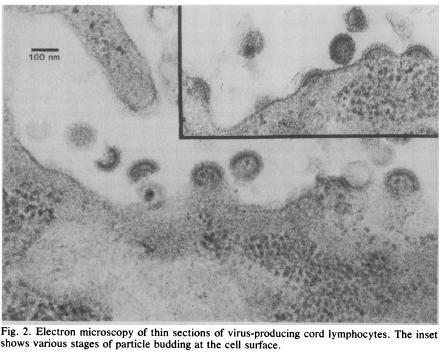{kind=link}
58. Perth Group translation of Extraits de l’interview de Charles Dauguet (Décembre 2005 – Paris). 2005. In December 2005 Djamel Tahi interviewed Charles Dauguet, the Pasteur Institute electron microscopist and one of the co-authors of the 1983 Montagnier paper. Dauguet stated that examination of density gradient material was performed because this procedure concentrates retrovirus particles. However, over a period of 15 days he could not find any retroviral- like particles in the density gradient "purified virus". He only found cellular debris. Asked if he also examined controls Dauguet replied “The samples on which I worked were from infected cultures”. 59. Gelderblom HR, Özel M, Hausmann EHS, Winkel T, Pauli G, Koch MA. Fine Structure of Human Immunodeficiency Virus (HIV), Immunolocalization of Structural Proteins and Virus-Cell Relation. Micron Microscopica 1988. 19:41-60. 60. Nermut M, Steven A. Animal virus structure. Elsevier Science, 1987. 61. Klatzmann D, Barré-Sinoussi F, Nugeyre MT, Danquet C, Vilmer E, Griscelli C, Brun-Veziret F, Rouzioux C, Gluckman JC, Chermann J-C. Selective tropism of lymphadenopathy associated virus (LAV) for helper-inducer T lymphocytes. Science 1984. 225:59-63. 62. Montagnier L, Gruest J, Chamaret S, Dauguet C, Axler C, Guetard D, Nugeyre MT, Barré- Sinoussi F, Chermann JC, Brunet JB, Klatzmann D, Gluckman JC. Adaption of lymphadenopathy associated virus (LAV) to replication in EBV-transformed B lymphoblastoid cell lines. Science 1984. 225:63-66. 63. Kuznetsov YG, Victoria JG, Robinson WE, Jr., McPherson A. Atomic force microscopy investigation of human immunodeficiency virus (HIV) and HIV-infected lymphocytes. J Virol 2003. 77:11896-11909. 64. The knob count has been claimed to be 80, 72, approximately 14, 0.5 (on average) and possibly zero. Kuznetsov and his colleagues wrote, "The clusters of gp120 [a constituent protein] do not form spikes on the surface of HIV as is commonly described in the literature. The clusters are hardly protrusions at all. We suggest that the spikes observed by negative-staining electron microscopy may be an artifact of the penetration of heavy metal stain between envelope proteins. Indeed, the term “spike” appears to have assumed a rather imprecise, possibly misleading definition, and might best be used with caution". 65. Montagnier L. Nobel Lecture. 2008. http://www.nobelprize.org/mediaplayer/index.php?id=1056 http://www.nobelprize.org/nobel_prizes/medicine/laureates/2008/montagni… 66. Papadopulos-Eleopulos E, Turner V, Papadimitriou J, Causer D. Are Montagnier's particles a retrovirus? 2008. http://www.theperthgroup.com/Nobel/MontagnierEMNobel.pdf 67. If Montagnier followed the custom of presenting one's best pictures then the taxonomy of the particles Montagnier showed in the one image at his Nobel lecture defies classification. http://www.theperthgroup.com/Nobel/MontagnierEMNobel.pdf 68. Gelderblom HR, Hausmann EH, Ozel M, Pauli G, Koch MA. Fine structure of human immunodeficiency virus (HIV) and immunolocalization of structural proteins. Virology 1987. 156:171- 176. 69. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Causer D. Factor VIII, HIV and AIDS in haemophiliacs: an analysis of their relationship. Genetica 1995. 95:25-50. http://www.theperthgroup.com/SCIPAPERS/HaemophiliaHIVAIDS.pdf 70. Hausmann EHS, Gelderblom HR, Clapham PR, Pauli G, Weiss RA. Detection of HIV envelope specific antibodies by immunoelectron microscopy and correlation with antibody titer and virus neutralizing activity. J Virol Methods 1987. 16:125-137. 71. Zhu P, Liu J, Bess J, Jr., Chertova E, Lifson JD, Grise H, Ofek GA, Taylor KA, Roux KH. Distribution and three-dimensional structure of AIDS virus envelope spikes. Nature 2006. 441:847- 852. 72. Brief Communications Arising from Zhu et al. 2006. Paper rejected by Nature. http://theperthgroup.com/LATEST/ZhuNatureRejected.doc 73. Chertova E, Bess Jr JW, Crise BJ, Sowder II RC, Schaden TM, Hilburn JM, Hoxie JA, Benveniste RE, Lifson JD, Henderson LE. Envelope glycoprotein incorporation, not shedding of surface envelope glycoprotein (gp120/SU), is the primary determinant of SU content of purified human immunodeficiency virus type 1 and simian immunodeficiency virus. J Virol 2002. 76:5315-5325.
62
74. Layne SP, Merges MJ, Dembo M, Spouge JL, Conley SR, Moore JP, Raina JL, Renz H, Gelderblom HR, Nara PL. Factors underlying spontaneous inactivation and susceptibility to neutralization of human immunodeficiency virus. Virology 1992. 189:695-714. 75. Burton DR. Structural biology: images from the surface of HIV. Nature 2006. 441:817-818. 76. Panem S. C Type Virus Expression in the Placenta. Curr Top Pathol 1979. 66:175-189. 77. Bader JP. Reproduction of RNA Tumor Viruses. Edited by Fraenkel-Conrat H, Wagne RR. New York, Plenum Press, 1975, pp. 253-331.
78. Dourmashkin RR, Bucher D, Oxford JS. Small virus-like particles bud from the cell membranes of normal as well as HIV-infected human lymphoid cells. J Med Virol 1993. 39:229-232. 79. Dourmashkin RR, O'Toole CM, Bucher D, Oxford JS. The presence of budding virus-like particles in human lymphoid cells used for HIV cultivation. Florence, 1991. 80. In 1993 Robert Dourmashkin reported the presence of retrovirus-like particles in human umbilical cord lymphocytes. "Electron microscopy (EM) of cell sections showed cell associated virus- like particles (VLP), 50-60 nm in diameter, budding from the membrane of human lymphoid cells in culture. The particles had an envelope continuous with the cell membrane and a dense core that almost filled the particle. Particles 70-80 nm in diameter with prominent external spikes were found in the culture medium by negative staining (medium-associated VLP). Cell-associated VLP were also present in cord lymphocytes, both on initial separation and after culture with or without foetal calf serum, and therefore were considered to be endogenous to the cells...VLP were observed in most of the lymphoid cell lines examined". 81. Popovic M, Sarin PS, Robert-Gurroff M, Kalyanaraman VS, Mann D, Minowada J, Gallo RC. Isolation and transmission of human retrovirus (human t-cell leukemia virus). Science 1983. 219:856- 859. 82. Hockley DJ, Wood RD, Jacobs JP. Electron Microscopy of Human Immunodeficiency Virus. J Gen Virol 1988. 69:2455-2469. 83. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Virus Challenge: Isolated facts about HIV-- A reply to Robin Weiss. Continuum 1996. 4:24-27. http://www.theperthgroup.com/CONTINUUM/VirusChallenge.pdf 84. Lecatsas G, Taylor MB. Pleomorphism in HTLV-III, the AIDS virus. S Afr Med J 1986. 69:793- 794. 85. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Causer D. Why no whole virus? Continuum 1997. 4:27-30. http://theperthgroup.com/CONTINUUM/WhyNoWholeVirus.pdf 86. O'Hara CJ, Groopman JE, Federman M. The ultrastructural and immunohistochemical demonstration of viral particles in lymph nodes from human immunodeficiency virus-related and non- human immunodeficiency virus-related lymphadenopathy syndromes. Hum Pathol 1988. 19:545-549. 87. Ho DD, Neumann AU, Perelson AS, Chen W, Leonard JM, Markowitz M. Rapid turnover of plasma virions and CD4 lymphocytes in HIV-1 infection. Nature 1995. 373:123-126. 88. Wei X, Ghosh SK, Taylor M, Johnson VA, Emini EA, Deutsch P, Lifson JD, Bonhoeffer S, Nowak MA, Hahn BH, Saag MS, Shaw GM. Viral dynamics in human immunodeficiency virus type 1 infection. Nature 1995. 373:117-122. 89. Piatak M, Jr., Saag MS, Yang LC, Clark SJ, Kappes JC, Luk KC, Hahn BH, Shaw GM, Lifson JD. High levels of HIV-1 in plasma during all stages of infection determined by competitive PCR. Science 1993. 259:1749-1754. 90. Hazelton PR, Gelderblom HR. Electron microscopy for rapid diagnosis of infectious agents in emergent situations. Emerg Infect Dis 2003. 9:294-303. 91. Braun DL, Kouyos R, Oberle C, Grube C, Joos B, Fellay J, McLaren PJ, Kuster H, Günthard HF. A novel acute retroviral syndrome severity score predicts the key surrogate markers for HIV-1 disease progression. PLoS ONE 2014. 9:e114111. 92. Moore J, Bergman J, Wainberg M. The AIDS denialists are still around. 2007. http://www.iasociety.org/Web/WebContent/File/Old/PDF/1293.pdf 93. Butler D. Medical journal under attack as dissenters seize AIDS platform. Nature 2003. 426:215. http://www.physics.smu.edu/pseudo/AIDS/DeclanButlerNov2003.pdf 94. Beard JW. Physical methods for the analysis of cells. Ann N Y Acad Sci 1957. 69:530-544. 95. Grafe A. A history of experimental virology. Heidelberg, Springer-Verlag, 1991, pp. 343.
63
96. Brun-Vezinet F, Barré-Sinoussi F, Saimot AG, Christol D, Rouzioux C, Klatzmann D, Rozenbaum W, Gluckmann JC, Chermann JC. Detection of IgG antibodies to lymphadenopathy- associatated virus in patients with AIDS or lymphadenopathy syndrome. Lancet 1984. I:1253-1256. 97. Matsiota P, Chamaret S, Montagnier L. Detection of Natural Autoantibodies in the serum of Anti-HIV Positive-Individuals. Annales de l'Institut Pasteur Immunologie 1987. 138:223-233.
98. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Phantom. Is a positive Western blot proof of HIV infection? Biotechnology (N Y) 1993. 11:696-707. http://leederville.net/links/EPENatBioTech1993.pdf 99. Zandman-Goddard G, Shoenfeld Y. HIV and autoimmunity. Autoimmun Rev 2002. 1:329-337. 100. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Causer D. The Isolation of HIV: Has it really been achieved? Continuum 1996. 4:1s-24s. http://www.theperthgroup.com/CONTINUUM/PapadopolousReallyAchieved1996.p…
101. Proof of "They [antibodies] know how to distinguish one molecule in one million” requires testing a large number of antibodies each against a panel of a million different antigens. There is no such proof. 102. Antibodies are proteins produced by the B cell lineage of lymphocytes. Each B-cell is unique in that it makes one and only one formula of antibody molecule. To produce that antibody in quantity the B-cells repeatedly divide to form a clone of the parental B-cell. Normally antibodies arise following exposure to foreign substances such as the proteins of a bacterium or virus. The antibodies bind to the substances which are given the general term antigens (from ANTIbody GENerating). Proteins are the most potent antigens and stimulate the production of many antibody molecules each of which reacts with one of the several, different, relatively small parts of the protein. These are called epitopes or antigenic determinants and the antibodies directed against each epitope are different molecules. Since the B cells that produce the antibodies only make one kind of antibody, many different clones of B cells are involved in an antibody response following exposure to any given protein. That is, the antibody response is polyclonal.
According to immunologist John Marchalonis “For many years, it was considered that a single antibody bound only the antigen [protein] to which it was raised...In fact, the concept arose that monoclonal antibodies must be monospecific [react with only one protein in the universe of proteins]. The immunological community was shocked to find that B cells [whose surfaces have antibody molecules attached to them] could be polyreactive in binding multiple antigens to their surface that were complex and ostensibly unrelated to one another”. Marchalonis described this behaviour as antibody "promiscuity". As long ago as 1969 the eminent Australian immunologist Sir Gustav Nossal wrote “An antibody molecule made following the injection of one antigen frequently can combine also with a second antigen...in other words, the antibody cross-reacts with the second antigen”.
In 2005 Paul Predki and his colleagues wrote: "The literature is replete with examples of crossreactive antibodies...the identity of cross-reactive proteins for the most part remains almost impossible to predict...Unrecognized, such cross-reactivity can have adverse consequences. The ability to assess and identify antibody cross-reactivity is an important but often inadequately addressed requirement for both research and clinical applications...”. The binding strength of a given antibody in a cross-reaction has been reported to be greater than for its “true” reaction, that is, for the protein against which the immune system produced the antibody in the first place. Predki documented this using a monoclonal antibody which reacted with 40 different protein antigens, binding to 16 of them more strongly than the antigen to which the antibody was raised. See Table 1 of Predki et al.
In 1997 Achim Kramer published data showing that a monoclonal antibody to the "specific" p24 “HIV” protein reacts with proteins found in bacteria, yeasts, amoebae, rabbits, monkeys and humans. The fungi include Candida albicans, the agent that causes one of the common AIDS indicator diseases. Nowadays a reaction between an anti-p24 antibody and proteins in a cell culture is considered proof of “HIV isolation”.
See:
64
Marchalonis JJ, Adelman MK, Robey IF, Schluter SF, Edmundson AB. Exquisite specificity and peptide epitope recognition promiscuity, properties shared by antibodies from sharks to humans. J Mol Recognit. 2001;14:110-121. Kramer A, Keitel T, Winkler K, Stocklein W, Hohne W, Schneider-Mergener J. Molecular basis for the binding promiscuity of an anti-p24 (HIV-1) monoclonal antibody. Cell. 12 1997;91:799-809.
Nossal GJV. Antibodies and Immunity. Harmondsworth, UK: Penguin Books Ltd; 1971. Predki PF, Mattoon D, Bangham R, Schweitzer B, Michaud G: Protein microarrays: a new tool for profiling antibody cross-reactivity, Hum Antibodies 2005, 14:7-15. 103. Marchalonis JJ, Adelman MK, Robey IF, Schluter SF, Edmundson AB. Exquisite specificity and peptide epitope recognition promiscuity, properties shared by antibodies from sharks to humans. J Mol Recognit 2001, 14:110-121. 104. Nossal GJV. Antibodies and Immunity. Harmondsworth, UK, Penguin Books Ltd, 1971, pp. 255. 105. Consider this experiment: A laboratory technician prepares two test-tubes each containing a colourless, aqueous solution of a compound whose identities are known only to him. He labels the tubes A and B and then asks a colleague to determine the identity of each compound. The scientist adds a few drops of the solution in tube B to tube A. Immediately dense clumps of a solid material (precipitate = proof of reaction) appear in tube A. The scientist then pronounces A is silver nitrate and B sodium chloride. This may be correct but the reaction is not proof because the same appearing precipitate would ensue if A = magnesium chloride and B = sodium hydroxide. Or A = barium chloride and B = sodium sulphate. And many other pairs of compounds. In these examples there are only two unknowns. In the Montagnier experiment there were many antigens (the As) and thousands of antibodies (the Bs) all of unknown origin and specificities. If one accepts Montagnier’s interpretation that A = p24 and B = antibody to p24 then one must accept there is no need for analytical chemistry or indeed analytical chemists. http://www.youtube.com/watch?v=DCl2VJUhjpY 106. The Australian immunologist Sir Gustav Nossal also places great faith in the ablility of antibodies to "recognise" the molecules with which they react. In his written statement to court at the 2006 Parenzee hearing Nossal wrote "...high affinity monoclonal antibodies are extensively used in research as razor-sharp and highly specific identifiers of various structures". These, essentially the same statements, are not supported by the evidence. To the contrary. As Marchalonis pointed out, in 1998 Van Regenmortel showed "there is no necessary correlation between affinity...and specificity because low affinity antibodies can show better discrimination among antigens than the high affinity binders". In 2005 Predki summarised the problem with "The literature is replete with examples of crossreactive antibodies. These examples have been identified both through the course of experimentation and through studies intentionally designed to characterize antibody specificity. Although in some cases researchers were able to identify potential sources for the observed cross- reactivity, the identity of cross-reactive proteins for the most part remains almost impossible to predict...Unrecognized, such cross-reactivity can have adverse consequences". The latter includes the futility of proving certain proteins as retroviral proteins based on their reactivity with unknown antibodies in serum of AIDS patients. Yet these are the basis of the belief in the existence of HIV and in the validity of tests to prove HIV infection. See: Van Regenmortel MH. From absolute to exquisite specificity. Reflections on the fuzzy nature of species, specificity and antigenic sites. Journal of Immunological Methods 216: 37-48, 1998. Predki PF, Mattoon D, Bangham R, Schweitzer B, Michaud G. Protein microarrays: a new tool for profiling antibody cross-reactivity. Hum Antibodies 14: 7-15, 2005.
107. Griner PF, Mayewski RJ, Mushlin AI. Selection and interpretation of diagnostic tests and procedures. Ann Intern Med 1981. 94:559-563. 108. Eleni Papadopulos-Eleopulos and Valendar Turner interviewed by Stuart Reid in the Australian Response to AIDS Oral History Project, 25 November 1993, National Library of Australia, ORAL TRC 2815/80. 1993.
109. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Has Gallo proven the role of HIV in AIDS? Emergency Medicine [Australia] 1993. 5:113-123. http://www.theperthgroup.com/SCIPAPERS/EPEGalloProveRoleHIVEmergMedOCR1…
65
110. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Is a positive Western blot proof of HIV infection? Biotechnology (N Y) 1993. 11:696-707. http://www.theperthgroup.com/SCIPAPERS/EPENatBioTech1993.pdf 111. Turner VF. The HIV Western blot. Med J Aust 1994. 160:807-808. http://www.theperthgroup.com/SCIPAPERS/VFTDax.pdf
112. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Stewart G, Causer D. HIV antibodies: further questions and a plea for clarification. Curr Med Res Opin 1997. 13:627-634. http://www.theperthgroup.com/SCIPAPERS/EPECurrMedResOpinHIVABFurther199… 113. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Causer D, Page BA. HIV antibody tests and viral load--more unanswered questions and a further plea for clarification. Curr Med Res Opin 1998. 14:185-186. http://www.theperthgroup.com/SCIPAPERS/EPECurrMedResOpinHIVAB&VLMore199…
114. Turner VF, McIntyre A. The Yin and Yang of HIV. NEXUS 1999. 6:29-36.
http://www.theperthgroup.com/POPPAPERS/yinyang.html
115. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Alfonso H, Page BAP, Causer D, Mhlongo S, Fiala C, Miller T, Brink A, Hodgkinson N. Mother to Child Transmission of HIV and its Prevention with AZT and Nevirapine. Perth, The Perth Group, 2001, pp. 204. 116. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Causer D. Global variation in the HIV Western blot. 2003. http://leederville.net/links/wbchart.doc
117. Turner VF. Detection of acute HIV infections. N Engl J Med 2005. 353:631-633; author reply 631-633. http://www.theperthgroup.com/SCIPAPERS/VFTNEJM.pdf 118. Papadopulos-Eleopulos E, Turner V, Papadimitriou J, Page B, Causer D. The "HIV" genome. 2008. www.theperthgroup.com/Nobel/MontagnierGenome.pdf
119. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Commentary on The Emperor's New Virus? 2012. http://theperthgroup.com/OTHER/ENVCommentary.pdf 120. Gamma globulins are a class of globulins, identified by their positions after serum protein electrophoresis. The most significant gamma globulins are immunoglobulins (antibodies), although some immunoglobulins are not gamma globulins, and some gamma globulins are not immunoglobulins. https://en.wikipedia.org/wiki/Gamma_globulin
121. McIntyre JA, Faulk WP. Redox-reactive autoantibodies: biochemistry, characterization, and specificities. Clin Rev Allergy Immunol 2009. 37:49-54. 122. McIntyre JA, Wagenknecht DR, Faulk WP. Autoantibodies unmasked by redox reactions. J Autoimmun 2005. 24:311-317.
123. McIntyre JA, Wagenknecht DR, Faulk WP. Redox-reactive autoantibodies: detection and physiological relevance. Autoimmun Rev 2006. 5:76-83. 124. Antibodies “recognising" HIV proteins (and presumably not "recognising" all others) is an example of the many anthropomorphisms* immunologists use to "explain" the immune system. Recognise means "to perceive something or someone as already known". Antibodies do not have memories. Antibodies are molecules which may or may not react with other molecules. Sodium chloride and silver nitrate react to produce silver chloride and sodium nitrate. Chemists do not claim this reaction is the result of molecular recognitions. From what Marcholanis and Kramer and many other scientists have reported, antibodies are unreliable witnesses: one and the same antibody may "recognise" many different molecules. Hence the identity of certain proteins as "HIV" and their use as antigens in the "HIV" antibody tests, is highly questionable.
*Further examples at www.abc.net.au/science/articles/2011/03/31/3177528.htm https://www.sciencedaily.com/releases/2014/11/141110124346.htm 125. The state of the War on Cancer is highlighted by the National Cancer Institute’s recruiting of cosmologist Professor Paul Davis in 2011 to head twelve new institutions to look at the cancer problem afresh. "As best he can remember, says Paul Davies, the telephone call that changed his professional life came some time in November 2007, as he was sitting in the small suite of offices that comprise his Beyond Center at Arizona State University (ASU) in Tempe...The caller — Anna Barker, then the deputy director of the US National Cancer Institute (NCI) in Bethesda, Maryland — explained that she needed his help in the ‘War on cancer’. Forty years into the government’s multibillion-dollar fight, said Barker, cancer survival rates had barely budged. The hope now was that physicists could bring some radical new ideas to the table, and she wanted Davies to give a keynote address at an
66
NCI workshop explaining how". Davies admitted "I don’t know anything about cancer” but Barker replied “We’re after fresh insights”. http://www.abc.net.au/radionational/programs/breakfast/physicist-versus…\ http://www.nature.com/news/2011/110601/full/474020a.html
126. It is not known how many individuals were diagnosed HIV-infected using the pre-1987 Western blot criteria of reactivity to p24 or p41 or both. Or how many, if any, were retested post-1987 and were still HIV-infected according to the newer criteria. Or how many studies on pathogenesis or epidemiology, if any, were revised in light of the newer criteria. 127. Brun-Vezinet F, Rouzioux C, Montagnier L, Chamaret S, Gruest J, Barré-Sinoussi F, Geroldi D, Chermann JC, McCormick J, Mitchell S, et al. Prevalence of antibodies to lymphadenopathy- associated retrovirus in African patients with AIDS. Science 1984, 226:453-456. 128. Saxinger WC, Levine PH, Dean AG, de The G, Lange-Wantzin G, Moghissi J, Laurent F, Hoh M, Sarngadharan MG, Gallo RC. Evidence for exposure to HTLV-III in Uganda before 1973. Science 1985, 227:1036-1038. 129. Morris L, Williamson C. Host and viral factors that impact on HIV-1 transmission and disease progression in South Africa. S Afr Med J 2001, 91:212-215. 130. USAID. Uganda and HIV/AIDS. U.S. Agency for International Development Population, Health and Nutrition Programs. HIV/AIDS Division. http://pdf.usaid.gov/pdf_docs/Pnacp184.pdf 131. WorldWatch Institute. Uganda on Track to Have World's Highest Population Growth. http://www.worldwatch.org/node/4525 132. Frazer IH, Mulhall BP. Second International Conference on the acquired immune deficiency syndrome. Med J Aust 1986. 145:524-529. 133. Clumeck N. Heterosexual Transmission of the Human Immunodeficiency Virus. Edited by Schinazi RF, Nahmias AJ. New York, Elsevier, 1988, pp. 160-162. 134. Serwadda D, Sewankambo NK, Carswell JW, Bayley AC, Tedder RS, Weiss RA, Mugerwa RD, Lwegaba A, Kirya GB, Downing RG, Clayden SA, Dagleish AG. Slim disease: A new disease in Uganda and its association with HTLV-III infection. Lancet 1985. ii:849-852. 135. Genesca J, Jett BW, Epstein JS, Shih JWK, Hewlett IK, Alter HJ. What do Western Blot indeterminate patterns for Human Immunodeficiency Virus mean in EIA-negative blood donors? Lancet 1989. ii:1023-1025. 136. Ranki A, Johansson E, Krohn K. Interpretation of antibodies reacting solely with human retroviral core proteins. N Engl J Med 1988. 318:448-449. 137. Agbalika F, Ferchal F, Garnier JP, Eugene M, Bedrossian J, Lagrange PH. False-positive HIV antigens related to emergence of a 25-30kD proteins detected in organ recipients. AIDS 1992. 6:959- 962. 138. Kashala O, Marlink R, Ilunga M, Diese M, Gormus B, Xu K, Mukeba P, Kasongo K, Essex M. Infection with human immunodeficiency virus type 1 (HIV-1) and human T cell lymphotropic viruses among leprosy patients and contacts: correlation between HIV-1 cross-reactivity and antibodies to lipoarabinomannan. J Infect Dis 1994. 169:296-304. 139. Lundberg GD. Serological diagnosis of human immunodeficiency virus infection by Western Blot testing. Journal of the American Medical Association 1988. 260:674-679. 140. Schupbach J, Jendis JB, Bron C, Boni J, Tomasik Z. False-positive HIV-1 virus cultures using whole blood. AIDS 1992. 6:1545-1546. 141. Coombs RW, Collier AC, Corey L. Plasma viraemia as an endpoint in evaluating the effectiveness of drugs against human immunodeficiency virus type-1 (HIV) infection: natural history of plasma viraemia and monitoring of antiretroviral therapy. Edited by Andrieu JM. Paris, John Libbey Eurotext, 1991, pp. 9-19. 142. Kozhemiakin LA, Bondarenko IG. Genomic instability and AIDS. Biokhimiia 1992. 57:1417- 1426. 143. Fauci AS, Lane HC. Chapter 189. Human Immunodeficiency Virus Disease: AIDS and Related Disorders. Edited by Longo DL, Fauci AS, Kasper DL, Hauser SL, Jameson JL, Loscalzo J. New York, NY, The McGraw-Hill Companies, 2012. 144. Strandstrom HV, Higgins JR, Mossie K, Theilen GH. Studies with canine sera that contain antibodies which recognize human immunodeficiency virus structural proteins. Cancer Res 1990. 50:5628s-5630s.
67
145. St Louis ME, Olivo N, Critchley S, Rauch KJ, White CR, Munn VP, Dondero Jr TJ. Methods of surveillance for HIV infection at US sentinel hospitals. Public Health Rep 1990. 105:140. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1580057/pdf/pubhealthrep001… http://theperthgroup.com/OTHER/St.LouisExceptions.pdf
146. St Louis ME, Rauch KJ, Petersen LR, Anderson JE, Schable CA, Dondero TJ. Seroprevalence rates of human immunodeficiency virus infection at sentinel hospitals in the United States. The Sentinel Hospital Surveillance Group. N Engl J Med 1990. 323:213-218. 147. Nossal G. Report for the Parenzee Hearing - Antibody Tests for HIV. 2006. http://www.tig.org.za/Parenzee_prosecution_transcripts/Nossal.pdf 148. Kramer A, Keitel T, Winkler K, Stocklein W, Hohne W, Schneider-Mergener J. Molecular basis for the binding promiscuity of an anti-p24 (HIV-1) monoclonal antibody. Cell 1997. 91:799-809. 149. Ranga U. The saga of the HIV controversy. Resonance 2009. 14:472-498. http://ns1.ias.ac.in/resonance/Volumes/14/05/0472-0498.pdf 150. Papadopulos-Eleopulos E. Between the lines – a critical analysis of Luc Montagnier's interview answers to Djamel Tahi. Continuum 1997. 5:35-45. http://www.altheal.org/continuum/Vol5no2.pdf http://theperthgroup.com/CONTINUUM/EPEReponseToDjamelTahiInterview.pdf 151. Connor EM, Sperling RS, Gelber R, Kiselev P, Scott G, O'Sullivan MJ, VanDyke R, Bey M, Shearer W, Jacobson RL, et al. Reduction of maternal-infant transmission of human immunodeficiency virus type 1 with zidovudine treatment. Pediatric AIDS Clinical Trials Group Protocol 076 Study Group. N Engl J Med 1994. 331:1173-1180. 152. Rubinstein E. The Untold Story of HUT78. Science 1990. 248:1499-1507. 153. According to Gallo’s criteria for "Detection and isolation", combinations of (2) and (3) or (2) and (4) or (3) and (4) are proof for the existence of a new and unique retrovirus HIV. However these include proofs based solely on RT or electron microscopic appearances; or without RT activity. One cannot have a retrovirus in the absence of reverse transcriptase. 154. Christie H. Interview with Dr. Robert Gallo July 1st Palexpo Conference Centre Geneva. New York, 1998. https://www.youtube.com/watch?v=CHxVXuCHkpU 155. That Gallo regards the Western blot antibody test proof of virus isolation (it "worked well ‘cos we could isolate the virus when we did it”) is confirmed in the 1984 Shaw et al Science paper which Gallo co-authored. In this paper Gallo and his colleagues wrote "We have previously been able to isolate HTLV-III [HIV] from peripheral blood or lymph node tissue from most patients with AIDS or ARC (12), in concordance with the 85 to 100 percent seropositivity for HTLV-III in these groups". In this paper's References and Notes, note 12 reads "From 88 to 100 percent of AIDS patients, approximately 85 to 90 percent of ARC patients, and in one series, 21 percent of healthy homo-sexual men possess serum antibodies to HTLV-III, whereas less than 1 percent of normal heterosexuals have these antibodies (3-5, 13-15)". References 3-4 are the third and fourth May 1984 Science papers of Gallo et al which are serological analyses of “HIV” and patients respectively. Reference 5 is another serological analysis published in Lancet by Safai et al. References 13-14 are "in preparation" and reference 15 is "unpublished data". Hence the papers cited as proof of isolation of HIV "from most patients with AIDS or ARC" are either papers that do not report virus isolation experiments, or are unpublished. Notably, Gallo does not cite his second Science paper in which, according to his data, "true isolation" of HIV was achieved from only 26/72 (36%) of his AIDS patients. 156. Crewdson J. Science Fictions - A scientific mystery, a massive cover-up, and the dark legacy of Robert Gallo. Boston: Little Brown and Company, 2003, pp. 670. 157. Gallo was not always averse to publishing EMs of density gradient material. In 1975 he published an electron micrograph to support his case for the existence of a retrovirus isolated from a patient with acute leukaemia. The UK retrovirologist Robin Weiss named the virus HL23V but as events unfolded HL23V was not a retrovirus or even a virus. And, as Figure 1 part A in the paper shows, whatever the nature of the banded material shown, even if a virus, it was obviously not pure. In Science Fictions John Crewdson writes, "HL23V wasn't a human virus after all, but a mélange of three animal viruses - a wooly monkey virus, a gibbon ape virus, and a baboon virus - jumbled up together in a retroviral cocktail". See:
68
Gallagher RE, Gallo RC. Type C RNA Tumor Virus Isolated from Cultured Human Acute Myelogenous Leukemia Cells. Science. 1975;187:350-353.
- Culliton BJ. Inside the Gallo Probe. Science 1990. 248:1494-1498.
- Wong-Staal F, Hahn B, Manzuri V, Colombini S, Franchini G, Gelmann EP, Gallo RC. A
survey of human leukemias for sequences of a human retrovirus. Nature 1983. 302:626-628. 160. Gluschankof P, Mondor I, Gelderblom HR, Sattentau QJ. Cell membrane vesicles are a major contaminant of gradient-enriched human immunodeficiency virus type-1 preparations. Virology 1997. 230:125-133. 161. The Gluschankof and Bess papers deal with the same problem: Contamination of density gradient "purified" HIV. Gluschankof's paper is entitled "Cell Membrane Vesicles Are a Major Contaminant of Gradient-Enriched Human Immunodeficiency Virus Type-1 Preparations" whereas Bess' title is “Microvesicles Are a Source of Contaminating Cellular Proteins Found in Purified HIV-1 Preparations”. Bess' abstract includes the sentence “Identification and quantitation of cellular proteins associated with HIV-1 particles are complicated by the presence of nonvirion-associated cellular proteins that copurify with virions” (emphasis added). The terms "Major Contaminant", "Contaminating Cellular Proteins" and “copurify” are incompatible with "Purified HIV-1 preparations". 162. Bess JW, Gorelick RJ, Bosche WJ, Henderson LE, Arthur LO. Microvesicles are a source of contaminating cellular proteins found in purified HIV-1 preparations. Virology 1997. 230:134-144. http://leederville.net/links/Bess.pdf 163. ExPASy ViralZone: The Lentivirus HIV-1 virion is "Enveloped, spherical to pleomorphic in shape, 80-100 nm in diameter". http://viralzone.expasy.org/all_by_species/71.html 164. The diameters of Bess' particles are problematic for another reason. Retrovirus particles have a density of 1.16 g/ml, a physical property used to advantage in their purification. The Bess "HIV" particles have an average diameter of 234 nM which makes their diameter 234/120 = 1.95 times larger than the upper defining limit of retrovirus particles. Since volume is proportional to the cube of diameter, the Bess particles must have a volume nearly eight times that of the defining limit of retrovirus particles. The ratio of the average diameters of the Bess to the Gluschankof particles is 234/140 = 1.67. This means the Bess particles must have a mass approximately 4.7 times greater than the mass of the Gluschankof particles – an unusual finding for one and the same virus. http://leederville.net/links/BessEM.doc 165. Cellular microvesicles are common cellular structures. Some are generated intracellularly (exosomes) and others are shed fragments of the cytoplasmic membrane. https://en.wikipedia.org/wiki/Microvesicles. The link below shows a liver cell containing many cellular microvesicles, some with nucleoids (nucleus-like structures). Their diameter is 80-100nm and some have "spike-like" surfaces. See: DW Fawcett "The Cell" Chapter 6 page 392. http://www.ascb.org/fawcetts-the-cell/. http://leederville.net/links/LiverVesicles.jpg 166. Cantin R, Diou J, Belanger D, Tremblay AM, Gilbert C. Discrimination between exosomes and HIV-1: purification of both vesicles from cell-free supernatants. 2008. The abstract to this paper includes "Although enveloped retroviruses bud from the cell surface of T lymphocytes, they use the endocytic pathway and the internal membrane of multivesicular bodies for their assembly and release from macrophages and dendritic cells (DCs). Exosomes, physiological nanoparticles produced by hematopoietic cells, egress from this same pathway and are similar to retroviruses in terms of size, density, the molecules they incorporate and their ability to activate immune cells. Retroviruses are therefore likely to contaminate in vitro preparations of exosomes and vice versa and sucrose gradients are inefficient at separating them". 167. Electrophoresis is a procedure that separates a mixture of proteins in solution. An aliquot of the mixture is placed at one end of a gel across which a circa 100 DC voltage is applied. The gel can be visualised as a molecular sieve that separates proteins according to their molecular weights. Under the influence of the electric field, proteins move through the gel – the lighter proteins moving faster. After several hours the proteins become separated, the voltage removed and the gel stained with a protein-specific dye. This reveals the relative positions of the proteins as a series of dark, horizontal lines/bands. Bands are thicker lines and the darker the lines/bands the greater the
69
{kind=link}
concentration of protein at that postion in the gel. The molecular weight of each protein is approximated by comparing its position with the positions of proteins of known molecular weights (markers) electrophoresed at the same time in a parallel gel. One should note that molecular weights determined by electrophoresis are not precise. For example, a protein of molecular weight 24K might be measured as 25K, especially if the electrophoresis is performed on material obtained from different experiments in a different or even the same laboratory. 168. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. The Journal of cell biology 2013. 200:373-383. http://jcb.rupress.org/content/200/4/373.full.pdf+html 169. Henderson LE, Sowder R, Copeland TD. Direct Identification of Class II Histocompatibility DR Proteins in Preparations of Human T-Cell Lymphotropic Virus Type III. J Virol 1987. 61:629-632. 170. Pinter A, Honnen WJ, Tilley SA, Bona C, Zaghouani H, Gorny MK, Zolla-Pazner S. Oligomeric structure of gp41, the transmembrane protein of human immunodeficiency virus type 1. J Virol 1989. 63:2674-2679. 171. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Commentary on the Mulder paper in Lancet. 1993. http://theperthgroup.com/SOUTHAFRICA/MULDER.pdf 172. Papadopulos-Eleopulos E. A critical analysis of the evidence for the existence of HIV and the HIV antibody tests: Presentation via satellite to the XIIth International AIDS Conference, Geneva. 1998. http://theperthgroup.com/presentations.html 173. Turner VF. Where we have gone wrong? Continuum 1998. 5:38-44. http://theperthgroup.com/CONTINUUM/VFTContinuumGoneWrong1998.pdf http://www.virusmyth.com/aids/hiv/vtwrong.htm 174. Turner VF. Emergency physicians roles in managing HIV seroconversion illness: Take stock or take HAART? Emergency Medicine Australia 1999. 16:201-203. http://www.theperthgroup.com/SCIPAPERS/EMAHIVLetterandReply.pdf 175. Papadopulos-Eleopulos E, Turner V. Presentation to the Presidential Panel AIDS Advisory Meeting 3 & 4 July 2000 Johannesberg, South Africa. 2000. http://www.theperthgroup.com/PRESENTATIONS/pretoria2.doc 176. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Alfonso H, Page BAP, Causer D, Mhlongo S, Fiala C, Brink A. High rates of HIV seropositivity in Africa-alternative explanation. International Journal of STD & AIDS 2003. 14:426-427. http://www.theperthgroup.com/SCIPAPERS/PerthGroupIJSAJune3003.pdf 177. The statement "a DNA probe was hybridised with one or more unknown DNAs" often means the probe was added to the unknowns in order to see whether or not the probe will hybridise. 178. http://www.youtube.com/watch?v=SvjeCxVu2dI 179. Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular biology of the cell. New York, Garland Science, 2002. 180. Racaniello V. What is a virus? 2013. https://www.youtube.com/watch?v=-C0r_-1DufM 181. The Parenzee Case. In 2006/2007 Andre Chad Parenzee made an application for leave to appeal to his earlier conviction for endangering the lives of three women following "unprotected sexual intercourse...when he knew he was infected with the virus HIV". (R v PARENZEE [2007] SASC 143; 248 LSJS 99, https://jade.io/j/?a=outline&id=8353). http://www.tig.org.za/Transcript_Perth_Group_evidence.htm 182. Fredericks DN, Relman DA. Sequence-based identification of microbial pathogens: a reconsideration of Koch's postulates. Clin Microbiol Rev 1996. 9:18-33. 183. Felgner PL, Ringold GM. Cationic liposome-mediated tranfection. Nature 1989. 337:387-388. 184. Kitsis RN, Buttrick PM, McNally EM, Kaplan ML, Leinwald LA. Hormonal modulation of a gene injected into rat heart in vivo. Proc Natl Acad Sci U S A 1991. 88:4138-4142. 185. Wolff JA, Malone RW, Williams P, Chong W, Acsadi G, Jani A, Felgner PL. Direct gene transfer into mouse muscle in vivo. Science 1990. 247:1465-1468. 186. Fisher AG, Collati E, Ratner L, Gallo RC, Wong-Staal F. A molecular clone of HTLV-III with biological activity. Nature 1985. 316:262-265. 187. Maddox J. More on Gallo and Popovic. Nature 1992. 357:107-109. 188. Crewdson J. The Great AIDS Quest - Science under the microscope. Chicago, 1989. 189. Hirsch MS, Phillips SM, Solnik C. Activation of Leukemia Viruses by Graft-Versus-Host and Mixed Lymphocyte Reactions In Vitro. Proc Natl Acad Sci U S A 1972. 69:1069-1072.
70
190. Toyoshima K, Vogt PK. Enhancement and Inhibition of Avian Sarcoma Viruses by Polycations and Polyanions. Virology 1969. 38:414-426. 191. Culliton reported, "Why pool the viruses? Popovic reasoned that the failure to grow might be tied to the fact that none of the viruses individually was producing high concentrations of reverse transcriptase. Maybe if he dumped ten viruses into the same pot, the reverse transcriptase level would be enough to jolt one of them into action. "The logic behind that is really crazy," says one of the scientists who has commented on the Gallo documents for Science. "But there is no doubt that he did it"". If the individual levels of reverse transcriptase were insufficient to prove the existence of a retrovirus then the cells were not infected with a retrovirus. Mixing ten (or even a thousand) together does not alter this fact.
192. Gillespie D, Marshall S, Gallo RC. RNA of RNA tumor viruses contains poly A. Nature: New biology 1972. 236:227-231. 193. This is not the first time virologists have been misled by a chemical "surrogate marker" for retroviruses. In the 1920s a molecule, adenosine triphosphate (ATP) was discovered in biological tissue. This now well known molecule is split into adenosine diphosphate (ADP) and phosphate by an enzyme called adenosine triphosphatase (ATPase). In the 1950s ATPase was found to be “associated” with a retrovirus. “The ATPase was considered so consistent a component of virions” that is, the virus particles, that it was used to both detect and quantify the number of virus particles. However when it was realized that ATP and ATPases were found in all cells and that its presence in the oncovirus particles depended “upon cell-specific, not virus-specific factors”, use of this enzyme as a means of detecting retroviruses quietly disappeared.
See: Bader JP. Reproduction of RNA Tumor Viruses. In: Fraenkel-Conrat H, Wagne RR, eds. Comprehensive Virology. Vol 4. New York: Plenum Press; 1975:253-331.
194. Edmonds M. A history of poly A sequences: from formation to factors to function. Prog Nucleic Acid Res Mol Biol 2002. 71:285-389. 195. 143 RvPS. Testimony of Robert Gallo. 2006/7. http://www.tig.org.za/Parenzee_prosecution_transcripts/Gallo_complete.p…
196. Given that Gallo reportedly reviewed Montagnier's 1983 Science paper and wrote the abstract it is suprising Gallo did not know if Montagnier had purified the BRU virus. 197. Fassin D, Schneider H. The politics of AIDS in South Africa: beyond the controversies. BMJ 2003. 326:495-497. http://www.bmj.com/content/326/7387/495
198. Smith R. Milton and Galileo would back BMJ on free speech. Nature 2004. 427:287.
http://leederville.net/links/SmithNature2004.pdf
199. Davies S, Delamothe T. Revitalising rapid responses. Br Med J 2005. 330:1284.
http://bmj.bmjjournals.com/cgi/content/full/330/7503/1284
200. Judson H. The great betrayal: Fraud in science. Orlando, Harcourt, 2004, pp. 463. 201. Gotzsche PC, Delamothe T, Godlee F, Lundh A. Adequacy of authors' replies to criticism raised in electronic letters to the editor: cohort study. BMJ 2010. 341:c3926. http://www.bmj.com/content/341/bmj.c3926.full.pdf 202. McClintock B. Nobel Lecture. 1983. http://www.nobelprize.org/nobel_prizes/medicine/laureates/1983/mcclinto… 203. Check E. Genome project turns up evolutionary surprises. Nature 2007. 447:760-761. http://www.nature.com/encode/about/encode-project 204. Birney E, et al. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature 2007. 447:799-816. 205. 206. Pearson H. Genetics: what is a gene? The idea of genes as beads on a DNA string is fast fading. Protein-coding sequences have no clear beginning or end and RNA is a key part of the information package. Nature 2006. 441:398-401. Science writer Helen Pearson published this three-page article in Nature on ENCODE entitled "WHAT IS A GENE?" In it she wrote, “The idea of genes as beads on a DNA string is fast fading. Protein- coding sequences have no clear beginning or end and RNA is a key part of the information package". She reported the problem of defining the term "gene", "Without a clear definition of a gene, life is also
71
difficult for bioinformaticians who want to use computer programs to spot landmark sequences in DNA that signal where one gene ends and the next begins. But reaching a consensus over the definition is virtually impossible, as Karen Eilbeck can attest. Eilbeck, who works at the University of California in Berkeley, is a coordinator of the Sequence Ontology Consortium. This defines labels for landmarks within genetic-sequence databases of organisms, such as the mouse and fly, so that the databases can be more easily compared. The consortium tries, for example, to decide whether a protein-coding sequence should always include the triplet of DNA bases that mark its end. Eilbeck says that it took 25 scientists the better part of two days to reach a definition of a gene that they could all work with. “We had several meetings that went on for hours and everyone screamed at each other,” she says. The group finally settled on a loose definition that could accommodate everyone’s demands".
207. Noble D. Evolution beyond neo-Darwinism: a new conceptual framework. The Journal of experimental biology 2015. 218:1273. http://jeb.biologists.org/content/jexbio/218/1/7.full.pdf 208. Papadopulos-Eleopulos E. The Role of myosin and actin in carcinogenesis: an hypothesis. Speculations in Science and Technology 1981. 4:39-44. http://leederville.net/links/SpecSciTech1981.pdf
209. Covello PS, Gray MW. RNA editing in plant mitochondria. Nature 1989. 341:662-666. 210. Eisen H. RNA editing: Who's on first? Cell 1988. 53:331-332. 211. Lamond AI. RNA editing and the mysterious undercover genes of trypansomatid mitochondria. Trends Biochem Sci 1988. 13:283-284. 212. Maas S. Base modification RNA editing: information recoding on the fly. Semin Cell Dev Biol 2012. 23:243. 213. Gommans WM, Mullen SP, Maas S. RNA editing: a driving force for adaptive evolution? Bioessays 2009. 31:1137-1145. 214. Arrigo P, Pulliero A. Effect of Environmental Chemical Stress on Nuclear Noncoding RNA Involved in Epigenetic Control. Biomed Res Int 2015. http://dx.doi.org/10.1155/2015/761703 215. Maas S. Posttranscriptional recoding by RNA editing. Advances in protein chemistry and structural biology 2012. 86:193-224. 216. Urnovitz HB, Tuite JJ, Higashida JM, Murphy WH. RNAs in the sera of Persian Gulf War veterans have segments homologous to chromosome 22q11.2. Clinical Diagnostic Laboratory Immunology 1999. 6:330-335. 217. Vaquero C, Sanceau J, Sondermeyer P, Falcoff R. Kinetics of messenger accumulation coding for IFN γ, related to modifications in the poly (A) RNA population of activated human lymphocytes. Nucleic Acids Res 1984. 12:2629-2640. 218. Kelleher CA, Wilkinson DA, Freeman JD, Mager DL, Gelfand EW. Expression of novel- transposon-containing mRNAs in human T cells. J Gen Virol 1996. 77:1101-1110. 219. Sharma B. Oxidative stress in HIV patients receiving antiretroviral therapy. Curr HIV Res 2014. 12:13-21. 220. Mandas A, Iorio EL, Congiu MG, Balestrieri C, Mereu A, Cau D, Dessì S, Curreli N. Oxidative imbalance in HIV-1 infected patients treated with antiretroviral therapy. BioMed Research International 2009. http://downloads.hindawi.com/journals/bmri/2009/749575.pdf 221. Day BJ, Lewis W. Oxidative stress in NRTI-induced toxicity: evidence from clinical experience and experiments in vitro and in vivo. Cardiovasc Toxicol 2004. 4:207-216. 222. Drug interference is another possibility. For example, the plasma concentrations of the reverse transcriptase inhibitor drugs that constitute HAART may inhibit the reverse transcription step in the PCR assay of "viral load". Or there may be other, presently unknown mechanism(s) involved. A relatively simple experiment would resolve such possibilities. 223. Papadopulos-Eleopulos E. A Mitotic Theory. J Theor Biol 1982. 96:741-758. http://www.theperthgroup.com/EPE/MitoticTheory.pdf http://leederville.net/links/JTBMitoticTheory1982.pdf http://goo.gl/uwp7CU 224. Garcea RL, Alberts BM. Comparative studies of histone acetylation in nucleosomes, nuclei, and intact cells. Evidence for special factors which modify acetylase action. J Biol Chem 1980. 255:11454-11463. http://www.ncbi.nlm.nih.gov/pubmed/7440548 225. Montagnier L. Apports de la recherche dans la lutte contra le Sida en Afrique. Edited by Pietteur M. Belgique, Collection Resurgence, 2004, pp. 224.
72
226. Vartanian JP, Meyerhans A, Henry M, Wain-Hobson S. High-resolution structure of an HIV-1 quasispecies: identification of novel coding sequences. AIDS 1992. 6:1095-1098. 227. Wain-Hobson S. Virological mayhem. Nature 1995. 373:102. 228. Perelson AS, Neumann AU, Markowitz M, Leonard JM, Ho DD. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science 1996. 271:1582-1586.
229. Becker JL, Hazan U, Nugeyre MT, Rey F, Spire B, Barré-Sinoussi F, Georges A, Teulieres L, Chermann JC. Infection of insect cell lines by the HIV virus, an agent of AIDS, and a demonstration of insects of African origin infected by this virus. C R Acad Sci III 1986. 303:303-306. 230. Webster AD, Dalgleish AG, Malkovsky M, Beattie R, Patterson S, Asherson GL, North M, Weiss RA. Isolation of retroviruses from two patients with "common variable" hypogammaglobulinaemia. Lancet 1986. 1:581-583.
231. Ciampolillo A, Mirakian R, Schulz T, Marini V, Buscema M, Pujol-Borrell R, Franco Bottazzo G. Retrovirus-Like Sequences In Graves'disease: Implications For Human Autoimmunity. The Lancet 1989. 333:1096-1100. 232. Horwitz MS, Boyce-Janino MT, Faras AJ. Novel human endogenous sequences related to human immunodeficiency virus type 1. J Virol 1992. 66:2170-2179. 233. The polymerase chain reaction. https://www.youtube.com/watch?v=0rQFnbcEsog https://www.youtube.com/watch?v=OK7_ReXhVaQ anscriptase coding sequences in human nucleic acids: relation to primate retroviruses. J Virol 1989. 63:64-75. 234. Shih A, Misra R, Rush MG. Detection of multiple, novel reverse transcriptase coding sequences in human nucleic acids: relation to primate retroviruses. J Virol 1989. 63:64-75. 235. Skloot R, Turpin B. The immortal life of Henrietta Lacks. 2010. 236. de Mendoza C, Holguin A, Soriano V. False positive for HIV using commercial viral load quantification assays. AIDS 1998. 12:2076-2077. 237. Rich JD, Merriman NA, Mylonakis E, Greenough TC, Flanigan TP, Mady BJ, Carpenter CCJ. Misdiagnosis of HIV infection by HIV-1 plasma viral load testing: A case series. Ann Intern Med 1999. 130:37-39. 238. Cooper. Expert witness report to the South Australian Criminal Court. 2006. http://www.tig.org.za/Parenzee_prosecution_transcripts/Cooper_complete… 239. McDonald P. Expert witness report to the South Australian Criminal Court. 2006. http://www.tig.org.za/Parenzee_prosecution_transcripts/McDonald_first_c… 240. Britten RJ. Divergence between samples of chimpanzee and human DNA sequences is 5%, counting indels. Proceedings of the National Academy of Sciences 2002. 99:13633-13635. 241. Innocenti P, Ottmann M, Morand P, Leclercq P, Seigneurin JM. HIV-1 in blood monocytes: frequency of detection of proviral DNA using PCR and comparison with the total CD4 count. AIDS Research & Human Retroviruses 1992. 8:261-268. 242. Michael NL, Chang G, Ehrenberg PK, Vahey MT, Redfield RR. HIV-1 proviral genotypes from the peripheral blood mononuclear cells of an infected patient are differentially represented in expressed sequences. J Acquir Immune Defic Syndr 1993. 6:1073-1085. 243. Meyerhans A, Cheynier R, Albert J, Seth M, Kwok S, Sninsky J, Morfeldt-Manson L, Asjo B, Wain-Hobson S. Temporal fluctuations in HIV quasispecies in vivo are not reflected by sequential HIV isolations. Cell 1989. 58:901-910. 244. ABC Radio Australia. The Science Show: How the body recognises viruses. 2014. http://www.abc.net.au/radionational/programs/scienceshow/how-the-body-r…- viruses/5690716#transcript 245. Owens DK, Holodniy M, Garber AM, Scott J, Sonnad S, Moses L, Kinosian B, Schwartz JS. Polymerase chain reaction for the diagnosis of HIV infection in adults. A meta-analysis with recommendations for clinical practice and study design. Ann Intern Med 1996. 124:803-815. 246. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Hedland-Thomas B, Causer D, Page B. Chart of the global variation in the criteria for a positive HIV Western blot. 1993. http://leederville.net/links/wbchart.doc 247. It is essential to understand the test parameters sensitivity and specificity. Sensitivity is a number, usually expressed as a percentage, indicating how often the test is positive given that a particular condition or disease is known to be present. For example, how many of 100 patients with
73
histolgically proven appendicitis have an elevated white blood cell count? The method of determining the presence of the condition, the test's gold standard, cannot be the test. The gold standard must be independent of the test. As another example, an ultrasound examination conducted at six weeks of pregnancy can serve as a gold standard for evaluating a blood test to diagnose pregnancy. If 99/100 women pregnant on ultrasound have a positive test then the test is 99% sensitive. Specificity is a more difficult concept because it is defined as a double negative. Specificity is the percentage of negative tests in a group of individuals who are known not to have the condition or disease. For example, if 99/100 women who are not pregnant on ultrasound have a negative blood test the test is 99% specific. The one non-pregnant woman with a positive test is a false-positive. This occurs for example in some gynaecological malignancies. The easy way to calculate the percent false-positives is to subtract the percent specificity from 100. Hence if the “HIV” PCR is 40% specific then 60% of individuals who are not infected will have a false-positive test. One cannot over-stress the requirement for using a superior reference test (the gold standard) to prove the condition or disease is present or absent. In the case of the “HIV” antibody tests there has never been a study documenting the test against HIV itself, despite the fact that proving HIV infection is the purpose of the test. The "HIV" antibody test is evaluated using either another antibody test (which is evaluating the test against itself), or by defining "HIV" infection as individuals who have AIDS. Neither method can prove that the antibodies that react in the test are caused by HIV. One must use HIV isolation/purification as the gold standard.
See: Griner PF, Mayewski RJ, Mushlin AI. Selection and interpretation of diagnostic tests and procedures. Ann Intern Med 1981 94:559-563.
248. One can judge the quality of PCR studies in general by the fact the authors of this review of PCR based their findings on 96 studies they deemed suitable for analysis from a collection of 379 studies which in turn they selected from 1735 titles judged to be potentially relevant. Forty-five of the studies analysed were published only as abstracts.
249. Defer C, Agut H, Garbarg-Chenon A, Moncany M, Morinet F, Vignon D, Mariotti M, Lefrere JJ. Multicentre quality control of polymerase chain reaction for detection of HIV DNA. AIDS 1992. 6:659- 663. 250. El Dib RP, Leeflang MM, Mathew JL, Almeida RA, Lewi DS, Kapoor A, Müller SS, Diaz RS. Nucleic acid amplification techniques (NAATs) for early diagnosis of HIV-1 and HIV-2 infections (Protocol). The Cochrane Library 2011. http://onlinelibrary.wiley.com/doi/10.1002/14651858.CD009184/pdf
251. The HIV experts use the antibody test as a gold standard for the PCR yet leading HIV experts assert that no gold standard exists for the HIV antibody tests. For example, according to Blattner "One difficulty in assaying the specificity and sensitivity of human retroviruses is the absence of a final gold standard". Antibody test kit manufacturers include similar disclaimers in their packet inserts. For example: "At present, there is no recognized standard for establishing the presence or absence of antibodies to HIV-1 and HIV-2 in human blood". Philip Mortimer, Director, Sexually Transmitted and Blood Borne Virus Laboratory, UK states, "Diagnosis of HIV infection is based almost entirely on detection of antibodies to HIV, but there can be misleading cross-reactions between HIV-1 antigens and antibodies formed against other antigens, and these may lead to false-positive reactions. Thus, it may be impossible to relate an antibody response specifically to HIV-1 infection". If there is no "final" or "recognized" gold standard how do HIV experts "relate an antibody response specifically to HIV-1 infection"?
See: Blattner WA. Retroviruses. Edited by Evans AS. New York, Plenum Medical Book Company, 1989, pp. 545-592. Abbott Axsym system (HIV-1/HIV-2). Abbott Laboratories, Diagnostics Division, 1998. Mortimer PP. The AIDS virus and the AIDS test. Medicine Internationale 1988, 56:2334-2339.
252. Branson BM, Owen SM, Wesolowski LG, Bennett B, Werner BG, Wroblewski KE, Pentella MA. Laboratory testing for the diagnosis of HIV infection: updated recommendations. Centers for
74
Disease Control and Prevention and Association of Public Health Laboratories 2014.
http://stacks.cdc.gov/view/cdc/23447 253. NEJM Journal Watch. It's time to dump the Western blot. 2012. http://blogs.jwatch.org/hiv-id- observations/index.php/its-time-to-dump-the-hiv-western-blot/2012/09/19/ 254. Rakowicz-Szulczynska EM, Jackson B, Szulczynska AM, Smith M. Human immunodeficiency virus type 1-like DNA sequences and immunoreactive viral particles with unique association with breast cancer. Clin Diagn Lab Immunol 1998. 5:645-653. 255. Kyriacou KC, Iacovou F, Adamou A, Hadjisavvas A, Rakowicz-Szulczynska EM. Immunohistochemical versus molecular detection of RAK antigens in breast cancer. Experimental and Molecular Pathology 2000. 69:27-36. 256. Rakowicz-Szulczynska EM, c Intosh M, David G, Morris P, Smith M. Novel Family of Gynecologic Cancer Antigens Detected by Anti-HIV Antibody. Infect Dis Obstet Gynecol 1994. 2:171- 178. 257. Rakowicz-Szulczynska EM, McIntosh DG, Smith ML. Giant syncytia and virus-like particles in ovarian carcinoma cells isolated from ascites fluid. Clin Diagn Lab Immunol 1999. 6:115-126. 258. Rakowicz-Szulczynska EM, Jackson B, Snyder W. Prostate, breast and gynecological cancer markers RAK with homology to HIV-1. Cancer Lett 1998. 124:213-223. 259. In the USA a woman is diagnosed with breast cancer every three minutes. Given the high sensitivity and specificity of Rakowicz’s data this may represent a far superior test for the early diagnosis of breast cancer than what is presently available. (The BRCA gene mutation is associated with 5-10% of breast cancers). From this point of view it does not matter what Rakowicz- Szulczynska's findings mean in terms of a retrovirus or the HIV theory of AIDS. If breast cancer tissue releases "HIV" DNA into the bloodstream, then detection of this DNA may prove to be a sensitive "liquid biopsy" for this disease. Since both sera from breast cancer patients and "HIV" PCR are readily available, it is puzzling that oncologists have not pursued this possibility. http://clinchem.aaccjnls.org/content/clinchem/61/1/112.full.pdf 260. Baxevanis A. Current topics in genome analysis 2012, Biological sequence analysis I. 2012. http://www.youtube.com/watch?v=Ud_6VpX5AgI&list=PLF09DBAA3E24C5068 261. Lesk A. Introduction to protein science: architecture, function, and genomics. Oxford University Press, 2010, pp. 480. 262. Kyte J, Doolittle RF. A simple method for displaying the hydropathic character of a protein. J Mol Biol 1982. 157:105-132. http://132.239.68.52/recentpubs/JMB1982.KyteRD.pdf 263. Romero Fernández-Bravo M. Contamination of genomic databases by HIV-1 and its possible consequences. A study in Bioinformatics. 2014. http://openaccess.uoc.edu/webapps/o2/handle/10609/31361 264. Romero's list of taxa include: Bienertia sinuspersici (a plant belonging to the Amaranth family), Bombus insularis (bees), Camellia sinensis (the camellia plant), an uncultured bacterial phylum from the human subgingival crevice, the fierce shrimpgoby coral reef fish, Culex pipiens quinquefasciatus (the southern house mosquito and vector of lymphatic filariasis and avian malaria), Dirofilaria immitis (a parasitic heartworm spread by mosquitos), Homo sapiens neanderthalensis, Homo sapiens chromosome 8, Human T-lymphotropic virus I, Leishmania major (a protozoan pathogen and cause of cutaneous leishmaniasis), Locusta migratoria (the migratory locust), Methyloversatilis universalis (a betaproteobacterium), Mus musculus (the house mouse), Neurospora intermedia (an Ascomycete fungus used in the production of the food staple oncom), Nicotiana tabacum (the tobacco plant), Oryza sativa Indica (long-grained rice), Phaseolus coccineus (runner bean), Plasmodium vivax (a protozoal parasite and cause of benign tertian malaria), Reticulitermes flavipes (eastern subterranean termite) gut metagenome, Sesamum indicum (the sesame plant), Setaria italica (the foxtail millet plant), Sorghum bicolor (sorghum), Streptomyces species (actinobacteria) and Zea mays (maize). 265. Merchant S, Wood DE, Salzberg SL. Unexpected cross-species contamination in genome sequencing projects. PeerJ 2014. 2:e675. 266. Contamination in Sequence Databases. http://www.ncbi.nlm.nih.gov/tools/vecscreen/contam/ 267. James Watson's genome sequenced at high speed. On April 16th 2008 Nature announced: "It took just four months, a handful of scientists and less than US$1.5 million to sequence the 6 billion base pairs of DNA pioneer James Watson". In two comments posted at the Nature online article,
75
Miguel Romero presents evidence that HIV-1 sequences are present in Watson's published genome.
http://www.nature.com/news/2008/080416/full/452788b.html http://www.nature.com/nature/journal/v452/n7189/full/nature06884.html 268. Romero Fernández-Bravo M. Readers comment on genome of Dr. James Watson. Nature 2014. 452:872-876. http://www.nature.com/nature/journal/v452/n7189/full/nature06884.html#c… http://www.nature.com/nature/journal/v452/n7189/full/nature06884.html#c… 269. Romero Fernández-Bravo M. Unexpected findings arising from Weisner et al. 2016. http://www.nature.com/nature/journal/v526/n7573/full/nature15258.html#c… 270. Romero Fernández-Bravo M. An unexpected finding arising from this study of the p53 tumor suppressor gene. 2016. http://www.plosgenetics.org/annotation/listThread.action?root=87413 271. Guidelines for the Use of Antiretroviral Agents in Pediatric HIV Infection. https://aidsinfo.nih.gov/guidelines/html/2/pediatric-arv-guidelines/55/…- infants-and-children 272. Papadopulos-Eleopulos E. Carcinogenesis. Med J Aust 1984. 140:180-181. http://theperthgroup.com/EPE/EPECarcinogenesisMJA1977.pdf 273. Papadopulos-Eleopulos E. Reappraisal of AIDS: Is the oxidation caused by the risk factors the primary cause? Med Hypotheses 1988. 25:151-162. http://www.theperthgroup.com/SCIPAPERS/EPEMedHyp1988.pdf 274. Papadopulos-Eleopulos E, Hedland-Thomas B, Causer DA, Dufty AP. An alternative explanation for the radiosensitization of AIDS patients. Int J Radiat Oncol Biol Phys 1989. 17:695-697. http://www.theperthgroup.com/SCIPAPERS/EPERadOncol1989.pdf 275. Papadopulos-Eleopulos E, Page BA, Causer D, Turner VF, Papadimitriou JM. Cancer and epigenetic reversion--the fundamental role of redox. Am J Pathol 2007. 171:1726-1727; author's reply p. 1727. http://theperthgroup.com/EPE/AMJLetterNov07.pdf 276. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Oxidative stress, HIV and AIDS. Res Immunol 1992. 143:145-148. http://www.theperthgroup.com/SCIPAPERS/EPEOxstressHIVAIDS.pdf 277. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Kaposi's sarcoma and HIV. Med Hypotheses 1992. 39:22-29. http://www.theperthgroup.com/SCIPAPERS/EPEMedHyp1992.pdf 278. Buhl R, Jaffe HA, Holroyd KJ, Wells FB, Mastrangeli A, Saltini C, Cantin AM, Crystal RG. Systemic glutathione deficiency in symptom-free HIV-seropositive individuals. Lancet 1989. 2:1294- 1298. 279. Eck HP, Gmunder H, Hartmann M, Petzoldt D, Daniel V, Droge W. Low concentrations of acid-soluble thiol (cysteine) in the blood plasma of HIV-1-infected patients. Biol Chem Hoppe Seyler 1989. 370:101-108. 280. Grek CL, Tew KD. Redox metabolism and malignancy. Current opinion in pharmacology 2010. 10:362-368. 281. Huber WW, Parzefall W. Thiols and the chemoprevention of cancer. Current opinion in pharmacology 2007. 7:404-409. 282. Papadopulos-Eleopulos E, Fox RA. Effects of dimethyl sulfoxide on cerebral arteries. Stroke 1987. 18:812. 283. Papadopulos-Eleopulos E, Knuckey N, Dufty A, Fox RA. Evidence that the redox state has a role in muscular contraction and relaxation. Physiol Chem Phys Med NMR 1985. 17:407-412. http://theperthgroup.com/EPE/MuscleRedox.pdf 284. Papadopulos-Eleopulos E, Knuckey N, Dufty A, Fox RA. Importance of the redox state in vasoconstriction induced by adrenaline and serotonin. Cardiovasc Res 1989. 23:662-665. http://theperthgroup.com/EPE/Serotonin.pdf 285. Pantel K, Alix-Panabieres C. Liquid biopsy: Potential and challenges. Molecular oncology 2016. http://www.ncbi.nlm.nih.gov/pubmed/26875532 286. Liquid cancer biopsy: the future of cancer detection? The Lancet Oncology 2016. 17:123. http://www.ncbi.nlm.nih.gov/pubmed/26868335 287. Salinas Sanchez AS, Martinez Sanchis C, Gimenez Bachs JM, Garcia Olmo DC. Liquid biopsy in cancer. Actas urológicas españolas 2016. 40:1-2. http://www.ncbi.nlm.nih.gov/pubmed/26296278
76
288. Ignatiadis M, Lee M, Jeffrey SS. Circulating Tumor Cells and Circulating Tumor DNA: Challenges and Opportunities on the Path to Clinical Utility. Clinical cancer research: an official journal of the American Association for Cancer Research 2015. 21:4786-4800. http://www.ncbi.nlm.nih.gov/pubmed/26527805 289. Goedert JJ, Sarngadharan MG, Biggar RJ, Weiss SH, Winn DM, Grossman RJ, Greene MH, Bodner AJ, Mann DL, Strong DM, Gallo RC, et al. Determinants of retrovirus (HTLV-III) antibody and immunodeficiency conditions in homosexual men. Lancet 1984. 2:711-716. 290. Stevens CE, Taylor PE, Zang EA, Morrison JM, Harley EJ, Rodriguez de Cordoba S, Bacino C, Ting RC, Bodner AJ, Sarngadharan MG, et al. Human T-cell lymphotropic virus type III infection in a cohort of homosexual men in New York City. Journal of the American Medical Association 1986. 255:2167-2172. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&do… 007789 291. Detels R, Jacobson L, Margolick J, Martinez-Maza O, Munoz A, Phair J, Rinaldo C, Wolinsky S. The multicenter AIDS cohort study, 1983 to.... Public Health 2012. 126:196-198. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3324261/pdf/nihms367040.pdf 292. Kingsley LA, Kaslow R, Rinaldo CR, Detre K, Odaka N, VanRaden M, Detels R, Polk BF, Chmiel J, Kelsey SF, Ostrow D, Visscher B. Risk factors for seroconversion to human immunodeficiency virus among male homosexuals. Lancet 1987. 329:345-348. 293. In the 1987 Multicenter AIDS cohort study (Kingsley et al) 2507 seronegative homosexual men were followed for six months. During this time seroconversions (new "infections") occurred in 95 men (3.8%), nine of whom claimed they did not practise passive anal sex. However, when further questioned, six of the nine admitted to practising passive anal sex in the months prior to the commencement of the study. Of the three remaining men the authors wrote, they “may reflect misclassification of men who actually did participate in receptive anal intercourse”. Even if these three men did not “reflect misclassification”, such small numbers lack the power to define an increased or even absolute risk greater than the zero risk observed in the 220 men in this study who did not practise receptive or insertive anal intercourse. 294. Caceres CF, van Griensven GJP. Male homosexual transmission of HIV-1. AIDS 1994. 8:1051-1061. 295. de Vincenzi I. A longitudinal study of human immunodeficiency virus transmission by heterosexual partners on Heterosexual Transmission of HIV. N Engl J Med 1994. 331:341-346. 296. Padian N, Marquis L, Francis DP, Anderson RE, Rutherford GW, O'Malley PM, Winkelstein W. Male-to-female transmission of human immunodeficiency virus. Journal of the American Medical Association 1987. 258:788-790. 297. Padian NS, Shiboski SC, Glass SO, Vittinghoff E. Heterosexual transmission of human immunodeficiency virus (HIV) in northern California: results from a ten-year study. Am J Epidemiol 1997. 146:350-357. 298. Hodgkinson N. The Failure of Contemporary Science: How a virus that never was deceived the world. London, Fourth Estate, 1996, pp. 420. 299. Marmor M, Friedman-Kien AE, Zolla-Pazner S, Stahl RE, Rubinstein P, Laubenstein L, William DC, Klein RJ, Spigland I. Kaposi's sarcoma in homosexual men. A seroepidemiologic case- control study. Ann Intern Med 1984. 100:809-815. 300. Marmor M, Friedman-Kien AE, Laubenstein L, Byrum RD, William DC, D'Onofrio S, Dubin N. Risk factors for Kaposi's sarcoma in homosexual men. Lancet 1982. i:1083-1087. 301. Amsterdam AIDS Cohort Studies. 2015. https://www.amsterdamcohortstudies.org/acsc/index.asp 302. Phair J, Jacobson L, Detals R, Rinaldo C, Saah A, Schrager L, Muñoz A. Acquired Immune Deficiency Syndrome Occuring Within 5 Years of Infection with Human Immunodeficiency Virus Type- 1: The Multicenter AIDS Cohort Study. J Acquir Immune Defic Syndr 1992. 5:490-496. 303. Nicholson JK, McDougal JS, Jaffe HW, Spira TJ, Kennedy MS, Jones BM, Darrow WW, Morgan M, Hubbard M. Exposure to human T-lymphotropic virus type III/lymphadenopathy-associated virus and immunologic abnormalities in asymptomatic homosexual men. Ann Intern Med 1985. 103:37-42.
77
304. Papadopulos-Eleopulos E. Looking back on the oxidative stress theory of AIDS. Continuum 1998. 5:30-35. http://theperthgroup.com/CONTINUUM/LookingBackContinuum.pdf 305. Reid BL. The causation of cervical cancer. Part I: A general review. Clin Obstet Gynaecol 1985. 12:1-18. http://www.ncbi.nlm.nih.gov/pubmed/3888478
306. Reid BL, French PW, Singer A, Hagan BE, Coppleson M. Sperm basic proteins in cervical carcinogenesis: Correlation with socioeconomic class. Lancet 1978. ii:60-62. 307. Stein-Werblowsky R. On the etiology of testicular tumors. An experimental study. Eur Urol 1978. 4:57-59. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&do… 27223
- Voeller B, Reinisch JM, Gottlieb M. AIDS and Sex. New York, Oxford University Press, 1990.
- Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, Kazzaz Z, Bornstein E,
Lambotte O, Altmann D, Blazar BR, Rodriguez B, Teixeira-Johnson L, Landay A, Martin JN, Hecht FM, Picker LJ, Lederman MM, Deeks SG, Douek DC. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med 2006. 12:1365-1371. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&do… 7115046
310. Marchetti G, Cozzi-Lepri A, Merlini E, Bellistrì GM, Castagna A, Galli M, Verucchi G, Antinori A, Costantini A, Giacometti A. Microbial translocation predicts disease progression of HIV-infected antiretroviral-naive patients with high CD4+ cell count. AIDS 2011. 25:1385-1394. 311. Vujkovic-Cvijin I, Dunham RM, Iwai S, Maher MC, Albright RG, Broadhurst MJ, Hernandez RD, Lederman MM, Huang Y, Somsouk M. Dysbiosis of the gut microbiota is associated with HIV disease progression and tryptophan catabolism. Science translational medicine 2013. 5:193ra191. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4094294/pdf/nihms587355.pdf
312. Daling JR, Weiss NS, Hislop TG, Maden C, Coates RJ, Sherman KJ, Ashley RL, Beagrie M, Ryan JA, Corey L. Sexual practices, sexually transmitted diseases, and the incidence of anal cancer. N Engl J Med 1987. 317:973-977. 313. Paglia C. " I am a celebrator of decadence." Michael Hattersley interviews the author of Vamps & Tramps. Harvard gay & lesbian review 1998. 5:11-14.
314. Christie H. Counterculture. Continuum 1996. 4:18-21.
http://www.altheal.org/continuum/Vol4no2.pdf
315. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Causer D. AIDS - Sexually transmitted or sexually acquired? 2010. http://www.theperthgroup.com/HIVEXIST/TPGSexTransMHNote.pdf 316. AVERT2. History of HIV and AIDS overview. http://www.avert.org/professionals/history-hiv- aids/overview
317. Jourjy J, Dahl K, Huesgen E. Antiretroviral Treatment Efficacy and Safety in Older HIV‐ Infected Adults. Pharmacotherapy: The Journal of Human Pharmacology and Drug Therapy 2015. 35:1140-1151. 318. Awards of Courage Joseph Sonnabend, M.D. Honoring with Pride 2000 Honoree. 2000. "Thanks to the new drugs we have, there are indeed people alive today who would otherwise be gone, and that’s a wonderful blessing. But I wish we knew how to use these drugs best. The only thing we know for sure is that very sick people will be helped — beyond any doubt they are really helped. What we don’t know is whether people who are asymptomatic ought to be taking drugs for 10 years — we just don’t know what that’s going to do to you. And if we were in a saner situation, we would say, ‘We need an answer, the only way to get an answer is to do a trial, so let’s do a trial.’ There is little hope that this will happen, but I never lose any opportunity to keep that hope alive". http://www.amfar.org/in-the-spotlight/awards-of-courage/2000-honoring-w…- sonnabend,-m-d-/
319. AIDSTruthTeam. The AIDSTruth: Our work is done. 2015.
https://www.aidstruth.org/2015/08/10/aidstruth-our-work-is-done/
320. If HIV was proven to be the cause of AIDS in 1984 and by 1987 there was no reasonable doubt, why was the Pasteur Institute inviting debate and manuscripts for publication in 1990, and accepting such papers until 1992? And why in January 1995 did John Maddox, the editor of Nature, decide, "Despite this journal's severe line, some months ago, on Duesberg's right of reply to critics of
78
his position, it is now in the general interest that his and his associates' views on the new developments should be made public...That will be eagerly awaited and will be published with the usual provisos". Maddox further wrote that the "new developments" led to "one important question [which] stands out like a sore thumb: why, after more than a decade of research, has it only now emerged that the response of the immune system to infection by HIV is hyperactivity rather than the opposite?". See: Maddox J. Duesberg and the new view of HIV. Nature 1995. 373:189.
- Doherty P. The Knowledge Wars. Melbourne University Press, 2015.
- Andreoni M, Marcotullio S, Puro V, De Carli G, Tambussi G, Nozza S, Gori A, Rusconi S,
Santoro M, Clementi M. An update on integrase inhibitors: new opportunities for a personalized therapy? The NEXTaim Project. The new microbiologica 2015. 38:443-490. 323. In June 2015 a paper in The Journal of Infectious Diseases reported a 30 year study of 20,858 HIV positive patients from San Francisco. The aim was to examine whether HAART “was associated with a better survival after a diagnosis of AIDS-defining opportunistic illnesses (AIDS- OIs) and how survival differed by AIDS-OI”. The “Overall 5-year survival probability increased from 7% in 1981–1986 to 65% in 1997–2012”. The ScienceDaily summary of this paper included “Thirty years of AIDS data highlight survival gains, room for improvement... “Although treatment advances have dramatically reduced deaths from opportunistic infections related to AIDS, a new study d rawing on 30 years of data from more than 20,000 patients in San Francisco suggests there is still ample room to improve. About a third--35 percent--of AIDS patients diagnosed with their first opportunistic infection from 1997 to 2012 in that city died within five years”. In other words, the five year survival with HAART is similar to that of lymphomas, melanoma and cancers of the larynx, uterus, cervix, prostate and breast. As Powel Kazanjian writes in The AIDS pandemic in historic perspective, “although HIV is not an automatic death sentence as it was in the early phase of the epidemic, it still remains a dangerous disease. Compared with an uninfected person, in fact, the lifespan of an HIV-infected person is shorter by approximately ten years on average. ART [antiretroviral therapy] has transformed HIV into a chronic disease that is not necessarily an easy one to live with”. See: Djawe K, Buchacz K, Hsu L, Chen M-J, Selik RM, Rose C, Williams T, Brooks JT, Schwarcz S. Mortality Risk After AIDS-Defining Opportunistic Illness Among HIV-Infected Persons—San Francisco, 1981–2012. J Infect Dis 2015 jiv235. ScienceDaily. Infectious Diseases Society of America. "Thirty years of AIDS data highlight survival gains, room for improvement.". ScienceDaily 2015. www.sciencedaily.com/releases/2015/06/150604084715.htm Quaresma M, Coleman MP, Rachet B. 40-year trends in an index of survival for all cancers combined and survival adjusted for age and sex for each cancer in England and Wales, 1971–2011: a population-based study. The Lancet 2015 385:1206-1218. Kazanjian P. The AIDS pandemic in historic perspective. J Hist Med Allied Sci 2014 69:351-382.
324. France B. Another kind of AIDS crisis. 2009. A striking number of HIV patients are living longer but getting older faster—showing early signs of dementia and bone weakness usually seen in the elderly. http://nymag.com/health/features/61740/ 325. In an extensive review article published in August 2013 from the Department of Cardiovascular Medicine, University of Oxford, it was reported that treatment with statins, by "redressing the myocardial nitroso-redox balance and reducing inflammation, has emerged as a potentially effective strategy for prevention of AF [atrial fibrillation]. Evidence indicates that statins prevent AF-induced electrical remodelling in animal models of atrial tachypacing and may reduce the new onset of AF after cardiac surgery". There is also the potential that statins reverse cardiac arrhythmias.
See: Dr. Ana-Catarina Pinho-Gomes, Prof. Svetlana Reilly, Prof. Ralph P. Brandes, and Barbara Casadei. Antioxidants & Redox Signaling. -Not available-, ahead of print. doi:10.1089/ars.2013.5542.
79
326. Bradley D. Why big pharma needs to learn the three 'R's. Nature Reviews Drug Discovery 2005. 4:446. 327. Roder C, Thomson MJ. Auranofin: Repurposing an Old Drug for a Golden New Age. Drugs in R&D 2015. 15:13-20.
328. Johansen LM, DeWald LE, Shoemaker CJ, Hoffstrom BG, Lear-Rooney CM, Stossel A, Nelson E, Delos SE, Simmons JA, Grenier JM, Pierce LT, Pajouhesh H, Lehar J, Hensley LE, Glass PJ, White JM, Olinger GG. A screen of approved drugs and molecular probes identifies therapeutics with anti-Ebola virus activity. Science Translational Medicine 2015. 7:290ra289. doi: 10.1126/scitranslmed.aaa5597 329. Cheng H, Lear-Rooney CM, Johansen L, Varhegyi E, Chen ZW, Olinger GG, Rong L. Inhibition of Ebola and Marburg viral entry by G protein-coupled receptor antagonists. J Virol 2015. 89:9932-9938. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4577884/pdf/zjv9932.pdf 330. Pantziarka P, Bouche G, Meheus L, Sukhatme V, Sukhatme VP, Vikas P. The repurposing drugs in oncology (ReDO) project. 2014. 331. ScienceDaily. Researchers say it's time to consider propranolol as an anti-cancer drug. 2016. www.sciencedaily.com/releases/2016/10/161013125954.htm 332. Mastrolorenzo A, Rusconi S, Scozzafava A, Barbaro G, Supuran CT. Inhibitors of HIV-1 protease: current state of the art 10 years after their introduction. From antiretroviral drugs to antifungal, antibacterial and antitumor agents based on aspartic protease inhibitors. Curr Med Chem 2007. 14:2734-2748. 333. Chow WA, Jiang C, Guan M. Anti-HIV drugs for cancer therapeutics: back to the future? The Lancet Oncology 2009. 10:61-71. 334. Anderson KW, Mast N, Hudgens JW, Lin JB, Turko IV, Pikuleva IA. Mapping of the Allosteric Site in Cholesterol Hydroxylase CYP46A1 for Efavirenz, a Drug That Stimulates Enzyme Activity. Journal of Biological Chemistry 2016. 291:11876-11886. 335. The fact that antiretroviral therapy is followed by an increase in the T4 cell count does not prove the drug responsible or has an "anti-HIV" effect. The original and still used "antiretroviral" drug, zidovudine (AZT), raises the T4 cell count in people who are free from HIV. This has been documented in individuals such as laboratory workers accidently "exposed to HIV" and then given "prophylactic AZT" for a month but who were initially and subsequently HIV negative. Jay Levy reported such cases and wrote "Most important, the cellular response [raised T4 count] that we observed to the treatment challenges current interpretations of the effect of AZT (and perhaps other antiviral drugs) on the number of lymphocytes in treated patients". It also shows the pitfalls of using "surrogate" markers. If AZT acts this way in non-infected indviduals its action in infected individuals may have nothing to do with effects on HIV. One should note that AZT is a component of some highly active antiretroviral therapies (HAART). See: Levy JA, Ramachadran B, Barker E, Guthrie J, Elbeik T. Plasma viral load, CD4+ cell count and HIV-1 production by cells. Science 1996 271:670-671. Milazzo L, Vaira LM, Cremoni L. CD4+ lymphocyte count variations in HIV-negative subjects treated with zidovudine. AIDS 1996 10:1444-1445.
336. Danel C, Moh R, Gabillard D, Badje A, Le Carrou J, Ouassa T, Ouattara E, Anzian A, Ntakpé J, Minga A. A Trial of Early Antiretrovirals and Isoniazid Preventive Therapy in Africa. The New England journal of medicine 2015. 373:808. 337. CDC. HIV/AIDS surveillance report. 1997. http://www.cdc.gov/hiv/pdf/library/reports/surveillance/cdc-hiv-surveil… 338. CDC. Epidemiological aspects of the current outbreak of Kaposi's sarcoma and opportunistic infections. N Engl J Med 1982. 306:248-252.
339. CDC. Kaposi's sarcoma and Pneumocystis pneumonia among homosexual men-New York City and California. Morbidity and Mortality Weekly Reports 1981. 30:305-307. 340. CDC. Update on Kaposi's Sarcoma and Opportunistic Infections in Previously Healthy Persons-United States. Morbidity and Mortality Weekly Reports 1982. 31:294-301.
80
341. Levine PH. The acquired immunodeficiency syndrome in persons with hemophilia. Ann Intern Med 1985. 103:723-726. 342. WHO. Acquired Immunodeficiency Syndrome (AIDS) WHO/CDC case definition for AIDS. Weekly Epidemiology Record 1986. 61:69-76.
343. Anonymous. CDC definition inadequate to handle growing cases, scientists complain. AIDS Alert 1987. July:120-121. 344. CDC. Revision of the CDC surveillance case definition for acquired immunodeficiency syndrome. Journal of the American Medical Association 1987. 258:1143-1154. https://www.cdc.gov/mmwr/pdf/other/mmsu3601.pdf
345. CDC. 1993 Revised Classification System for HIV Infection and Expanded Surveillance Case Definition for AIDS Among Adolescents and Adults. Morbidity and Mortality Weekly Reports 1992. 41:1-19. http://www.cdc.gov/mmwr/preview/mmwrhtml/00018871.htm 346. Brettle RP, Gore SM, Bird AG, McNeil AJ. Clinical and epidemiological implications fo the Centers for Disease Control/World Health Organization reclassifaction of AIDS cases. AIDS 1993. 7:531-539.
347. Roberts BD. HIV Antibody Testing Methods. Journal of Insurance Medicine 1994. 26:13-14.
https://aaimedicine.org/journal-of-insurance-medicine/jim/1994/026-01-0…
348. Clumeck N, Robert-Guroff M, Van De Perre P, Jennings A, Sibomana J, Demol P, Cran S, Gallo RC. Seroepidemiological studies of HTVL-III antibody prevalence among selected groups of heterosexual Africans. Journal of the American Medical Association 1985. 254:2599-2602. 349. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Bialy H. AIDS in Africa: Distinguishing fact and fiction. World Journal of Microbiology and Biotechnology 1995. 11:135-143. http://www.theperthgroup.com/SCIPAPERS/EPEWorldJMicro&BiotechOCR1995.pdf
350. Widy-Wirski R, Berkley S, Dowing R, Okware S, Recine U, Mugerwa R, Lwegaba A, Sempala S. Evaluation of the WHO clinical case defintion for AIDS in Uganda. Journal of the American Medical Association 1988. 260:3286-3289. 351. Fauci AS, Lane HC. Human Immunodeficiency Virus (HIV) Disease. In Harrinson's Principles of Internal Medicine. Eds. Isselbacher AS, Braunwald E, Wilson JD, Martin JB, Fauci AS, Kasper DL. McGraw Hill Inc. New York. 1993.
352. Dr. Claudia Kücherer. 2006. https://www.youtube.com/watch?v=BwgmzbnckII Time 31:06 353. Haverkos HW, Friedman-Kien AE, Drotman DP, Morgan WM. The changing incidence of Kaposi's sarcoma among patients with AIDS. J Am Acad Dermatol 1990. 22:1250-1253. 354. Drew W, Mills J, Hauer L, al. e. Declining prevalence of Kaposi's sarcoma in homosexual AIDS patients is paralleled by declining incidence of CMV infection. New Orleans, La, 1986.
355. Dore GJ, Li Y, Grulich AE, Hoy JF, Mallal SA, Mijch AM, French MA, Cooper DA, Kaldor JM. Declining incidence and later occurrence of Kaposi's sarcoma among persons with AIDS in Australia: the Australian AIDS cohort. AIDS 1996. 10:1401-1406. 356. Beral V, Peterman TA, Berkelman RL, Jaffe HW. Kaposi's sarcoma among persons with AIDS: a sexually transmitted infection? Lancet 1990. 335:123-128.
357. Bartlett JG. Ten years of HAART: Foundation for the future. 2006.
359. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Causer D, Alphonso H, Miller T. A critical analysis of the pharmacology of AZT and its use in AIDS. Curr Med Res Opin 1999. 15:1s-45s. http://www.theperthgroup.com/SCIPAPERS/AZTCMRO1999.pdf 360. Weber R, Ruppik M, Rickenbach M, Spoerri A, Furrer H, Battegay M, Cavassini M, Calmy A, Bernasconi E, Schmid P, Flepp M, Kowalska J, Ledergerber B, Swiss HIVCS. Decreasing mortality and changing patterns of causes of death in the Swiss HIV Cohort Study. HIV Med 2013. 14:195-207. http://www.ncbi.nlm.nih.gov/pubmed/22998068
361. Cohen J. Advances painted in shades of gray at a D.C. conference. Science 1997. 275:615- 616.
81
http://www.medscape.org/viewarticle/523119
358. Hleyhel M, Belot A, Bouvier AM, Tattevin P, Pacanowski J, Genet P, De Castro N, Berger JL, Dupont C, Lavole A. Trends in survival after cancer diagnosis among HIV‐infected individuals between 1992 and 2009. Results from the FHDH‐ANRS CO4 cohort. Int J Cancer 2015. 137:2443- 2453.
362. Dei-Cas E. Pneumocystis infections: the iceberg? Med Mycol 2000. 38 Suppl 1:23-32.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&do… 1204150 363. Heresi GP, Caceres E, Atkins JT, Reuben J, Doyle M. Pneumocystis carinii pneumonia in infants who were exposed to human immunodeficiency virus but were not infected: an exception to the AIDS surveillance case definition. Clin Infect Dis 1997. 25:739-740.
364. Jacobs JL, Libby DM, Winters RA, Gelmont DM, Fried ED, Hartman BJ, Laurence J. A Cluster of Pneumocystis Carinni Pneumonia in Adults without predisposing illnesses. N Engl J Med 1991. 324:246-250. 365. Stagno S, Pifer LL, Hughes WT, Brasfield DM, Tiller RE. Pneumocystis carinii pneumonitis in young immunocompetent infants. Pediatrics 1980. 66:56-62.
366. Stehbens WE. Oxidative stress in viral hepatitis and AIDS. Exp Mol Pathol 2004. 77:121-132.
http://www.ncbi.nlm.nih.gov/pubmed/15351235
367. Lauritsen J. Poison by prescription--The AZT story. New York, Asklepois Press, 1990. 368. Lauritsen J. The AIDS War: Propaganda, Profiteering and Genocide from the Medical- Industrial Complex. Asklepios, 1993. 369. Lauritsen JL. History of the Controversy. 2009. http://paganpressbooks.com/jpl/RA- SHORT.HTM
370. Anonymous. Concorde: MRC/ANRS randomised double-blind controlled trial of immediate and deferred zidovudine in symptom-free HIV infection. Concorde Coordinating Committee. Lancet 1994. 343:871-881. 371. Brink A. Debating AZT: Mbeki and the AIDS drug controversy. Open Books, 2000. http://www.tig.org.za/pdf-files/debating_azt.pdf. http://www.tig.org.za/
372. Richman DD, Andrews J. Results of continued monitoring of participants in the placebo- controlled trial of zidovudine for serious human immunodeficiency virus infection. Am J Med 1988. 85:208-213. 373. Richman DD, Fischl MA, Grieco MH, Gottlieb MS, Volberding PA, Laskin OL, Leedom JM, Groopman JE, Mildvan D, Hirsch MS, et al. The toxicity of azidothymidine (AZT) in the treatment of patients with AIDS and AIDS-related complex. A double-blind, placebo-controlled trial. N Engl J Med 1987. 317:192-197.
374. The AIDSTruth Team valedictory includes "The efficacy of antiretrovirals, starting with AZT in 1987, is unequivocal" but concedes "There were of course bumps in the early years of treatment. AZT was prescribed too early and in doses too high". Some of the "bumps" include the deaths of Rock Hudson, Arthur Ashe, Ryan White, Freddie Mercury, Rudolph Nureyev and Kimberly Bergalis. All died before the advent of HAART and while details of their treatments are largely unknown it is reasonable to assume all were treated with AZT monotherapy. During the era of AZT monotherapy the "doses too high" included 400 mg taken every four hours around the clock day and night. In the HAART era the recommended dose is 300 mg twice daily, that is, a 75% reduction from 2.4 g to 0.6 g. This plus the fact that many HAART regimes do not include AZT are expected to reduce the number of toxicities and hence "bumps". https://en.wikipedia.org/wiki/List_of_HIV-positive_people
375. Turner VF. Reducing agents and AIDS--Why are we waiting? Med J Aust 1990. 153:502.
http://www.theperthgroup.com/SCIPAPERS/RedAgent.pdf
376. Zaunders JJ, Cunningham PH, Kelleher AD, Kaufmann GR, Jaramillo AB, Wright R, Smith D, Grey P, Vizzard J, Carr A, Cooper DA. Potent antiretroviral therapy of primary human immunodeficiency virus type 1 (HIV-1) infection: Partial normalization of T lymphocyte subsets and limited reduction of HIV-1 DNA despite clearance of plasma viremia. J Infect Dis 1999. 180:320-329. 377. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Alfonso H, Page B, Causer D. Questions about results reported with potent antiretroviral therapy for human immunodeficiency virus type 1 infection. J Infect Dis 2000. 181:1518-1519. http://theperthgroup.com/SCIPAPERS/ZaundersLettersJID.pdf
378. Mikovits JA, Lohrey N, Schuloff R, Ruscetti F. Immune activation of latent HIV-1 expression in monocyte/macrophages. Florence, 1991. 379. Lundgren JD, et al. Initiation of Antiretroviral Therapy in Early Asymptomatic HIV Infection. N Engl J Med 2015. 373:795-807.
82
380. Munderi P, Walker A, Kityo C, Babiker A, Ssali F, Reid A, Darbyshire J, Grosskurth H, Mugyenyi P, Gibb D. Nevirapine/zidovudine/lamivudine has superior immunological and virological responses not reflected in clinical outcomes in a 48‐week randomized comparison with abacavir/zidovudine/lamivudine in HIV‐infected Ugandan adults with low CD4 cell counts. HIV Med 2010. 11:334-344.
381. Mills E, Kelly S, Bradley M, Mollon P, Cooper C, Nachega J. Antiretroviral effects on HIV‐1 RNA, CD4 cell count and progression to AIDS or death: a meta‐regression analysis. HIV Med 2008. 9:849-857. 382. According to the FDA "A surrogate marker "can be defined as “ ...a laboratory measurement or physical sign that is used in therapeutic trials as a substitute for a clinically meaningful endpoint that is a direct measure of how a patient feels, functions, or survives and is expected to predict the effect of the therapy"". http://www.ncbi.nlm.nih.gov/pmc/articles/PMC534924/
383. On four occasions beginning mid-2015 The Perth Group emailed Dr. Mills and his co-author Dr. Nachega: "You demonstrated that most HAART therapies offer high levels of CD4 and viral load control together with low rates of progression to AIDS/death. But you were unable to demonstrate a relationship between changes in surrogate markers and clinical events. This seems very surprising as the surrogate markers are not just ordinary surrogate markers. They are central to HIV pathogenesis. If a relationship between HIV viral load (HIV), CD4 and clinical outcomes could not be demonstrated, doesn't this at least suggest the cause of clinical syndrome may not be HIV? If there is no relationship between CD4 and clinical outcomes could this imply that the underlying cause of the AIDS indicator diseases may not be due to a decrease in CD4 cell numbers (immune deficiency)?" Neither author replied.
83